Erste Durchläufe mit der optomotorischen Wendereaktion
on Monday, June 15th, 2026 12:25 | by Krisztian Sabo
Als ich anfing, ging es bei mir gleich mit einem klassischen neurobiologischen Experiment los: der optomotorischen Wendereaktion.
Zunächst übte ich mich von Mitte bis Ende März an Wildtypen, den ganzen April über stand ich für die eigentlichen Experimente im Labor, um drei verschiedene Fliegenlinien zu testen:
- Linie 14: Das ist meine eingekreuzte Kontrolle, gebalanced über den TM6-Marker. Hier sollte das Nervensystem völlig intakt sein.
- Linie 5 & Linie 6: Das sind die reziproken Kreuzungen aus UAS-TnTG(211) und der Gal4-Treiberlinie R23G12. Hier wird Tetanustoxin (TnT) exprimiert, welches die synaptische Übertragung in den angesteuerten Neuronen blockiert, die Neurone werden also quasi stummgeschaltet.
Ergebnisse Linie 14
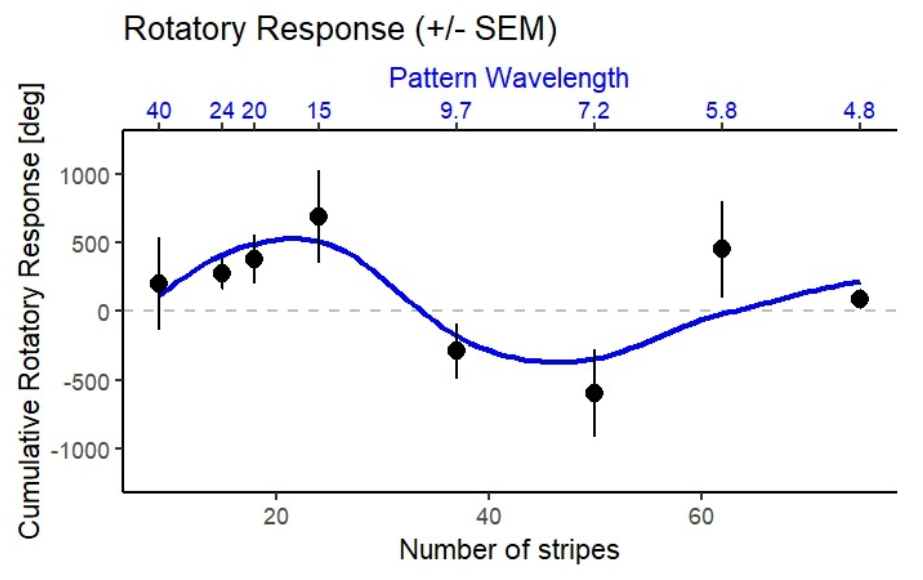
Die Kontrollfliegen zeigen eine starke, koordinierte Wendereaktion. Das System funktioniert also!
Da Linie 5 und Linie 6 genetisch reziproke Kreuzungen sind, habe ich die Daten Anfang Mai gepoolt, um eine solidere Datenbasis zu bekommen:
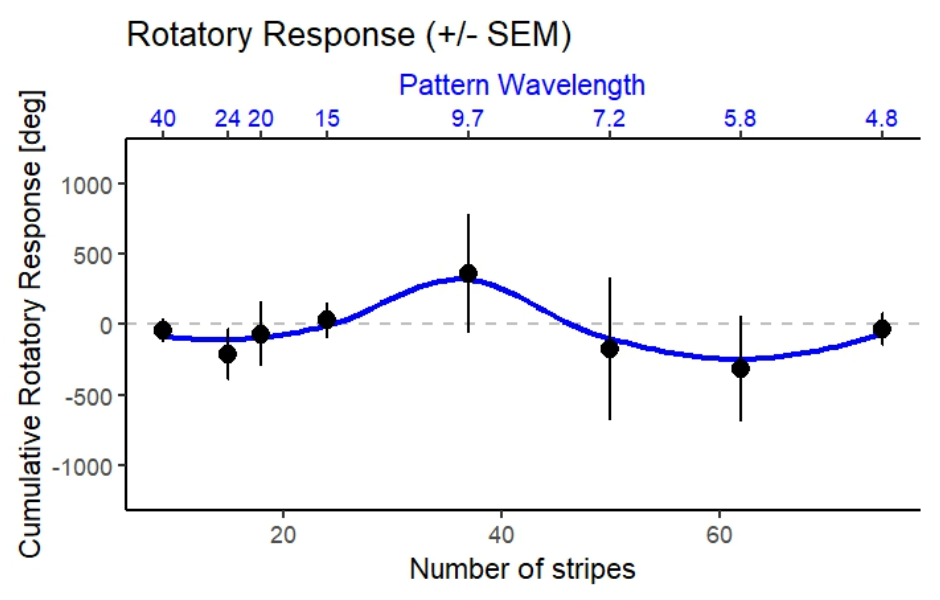
Im direkten Vergleich zur Kontrolle ist die optomotorische Wendereaktion bei den TnT-Fliegen massiv abgeflacht und über weite Teile der Reize fast vollständig inhibiert. Das Stummschalten der über R23G12 angesteuerten Neurone scheint die visuelle Folgereaktion also empfindlich zu stören oder gar ganz zu unterbrechen.

Category: Optomotor response, T4/T5 | No Comments
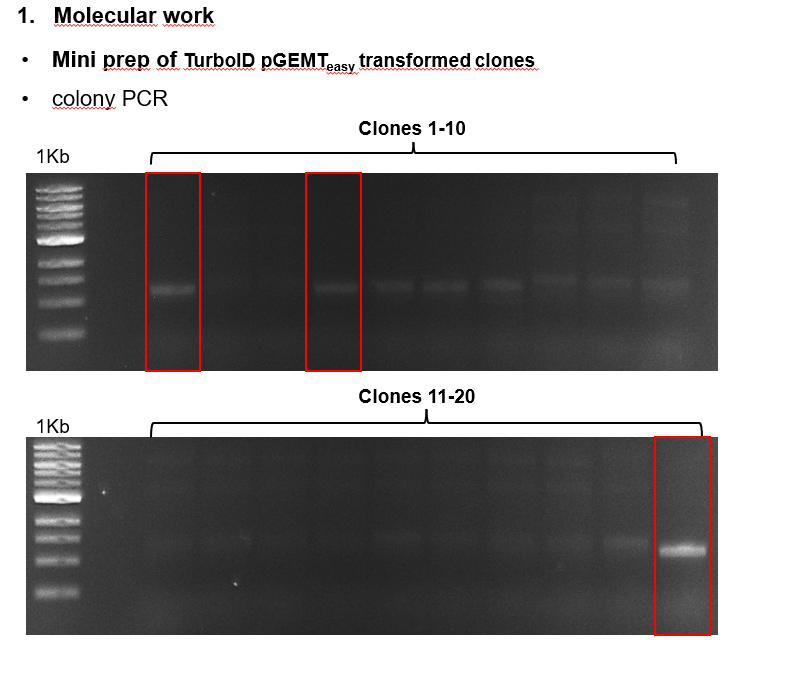
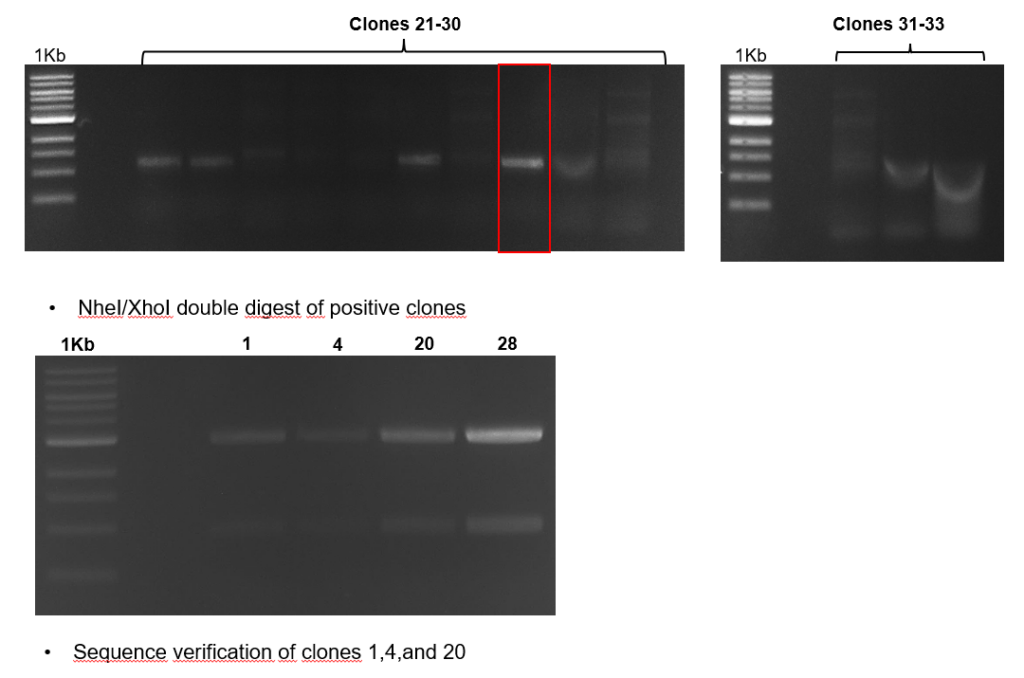



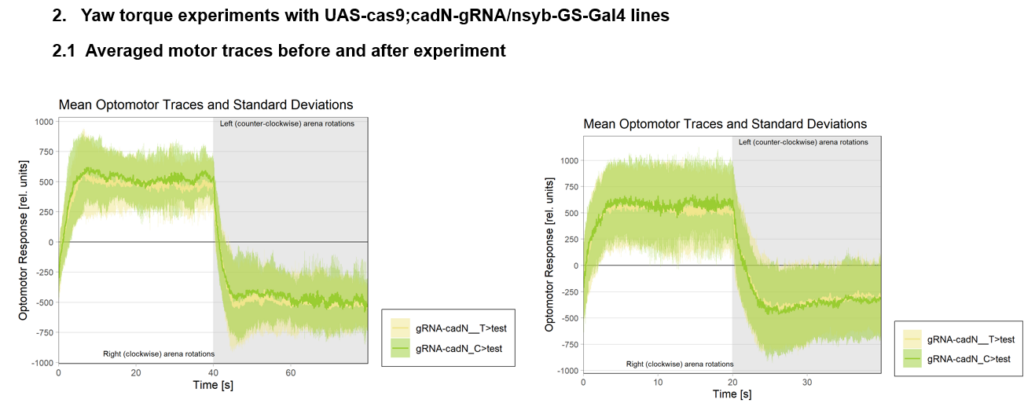
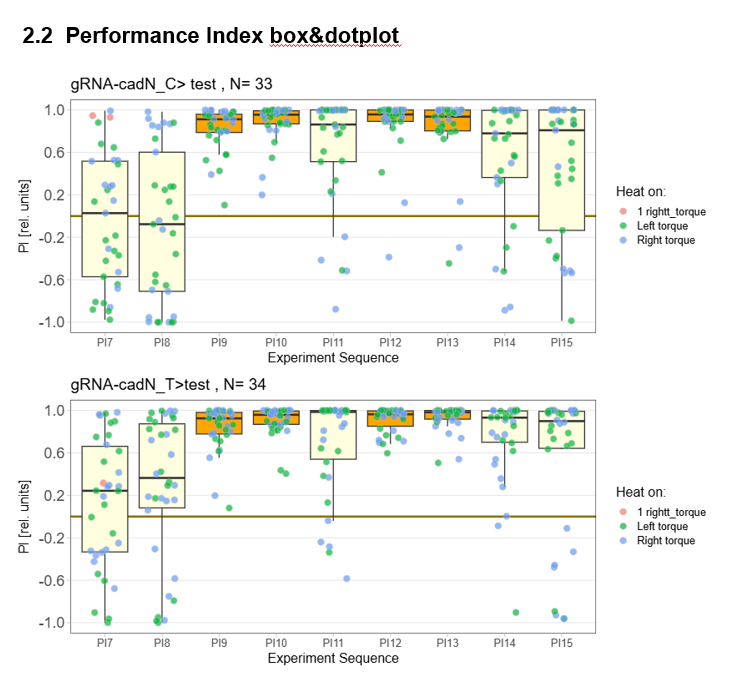
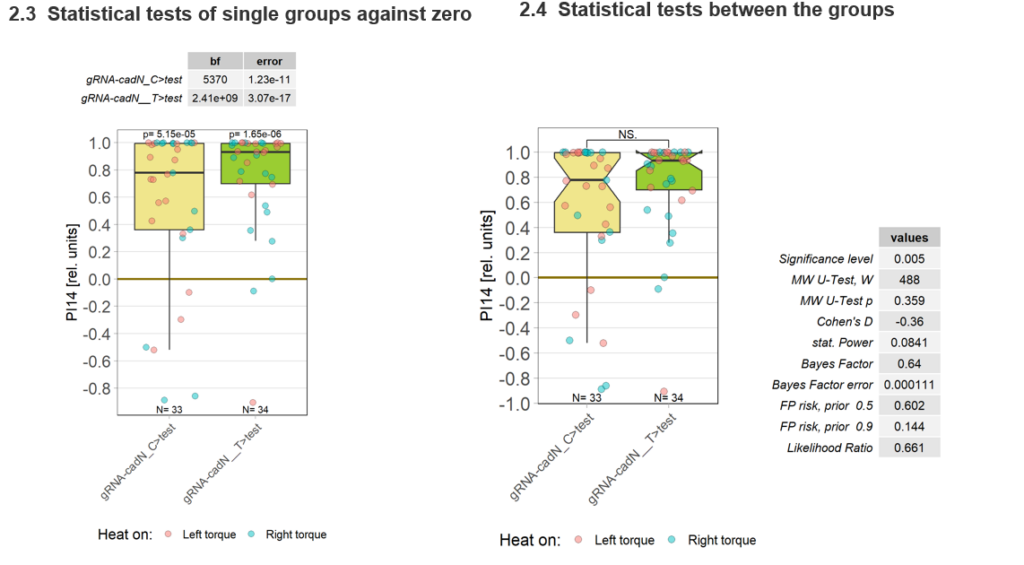
Torque measurements 21.02 & 22.02
on Monday, February 23rd, 2026 12:21 | by Fridrik Kjartansson
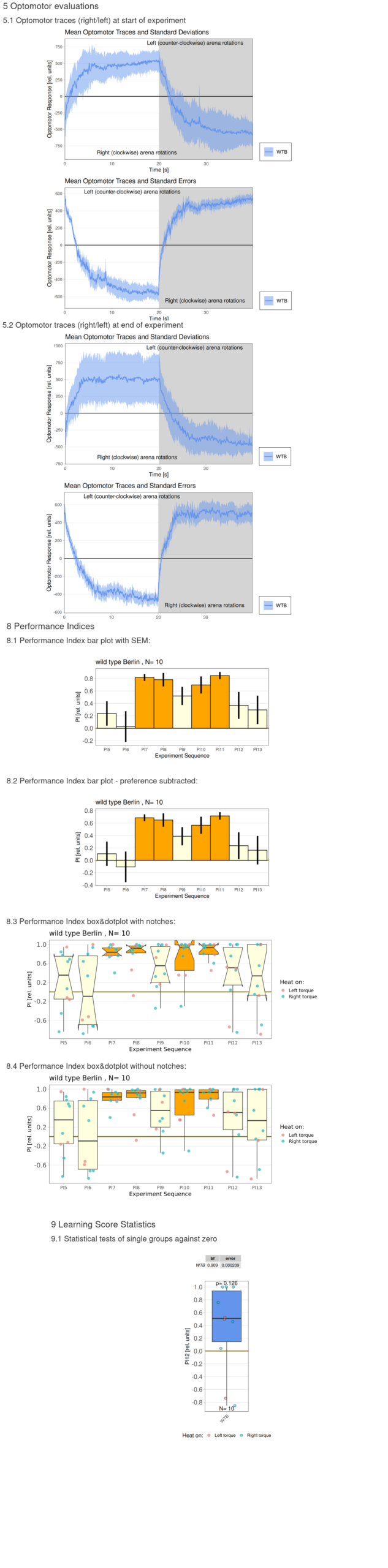
Category: flight, Habit formation, operant self-learning, Optomotor response | No Comments
Torque measurements 12.02.2026
on Friday, February 13th, 2026 12:38 | by Fridrik Kjartansson
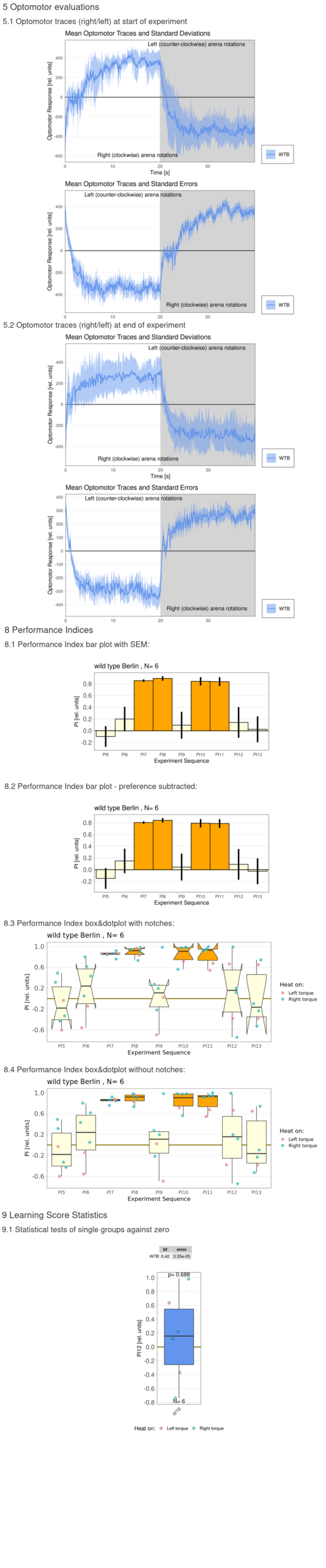
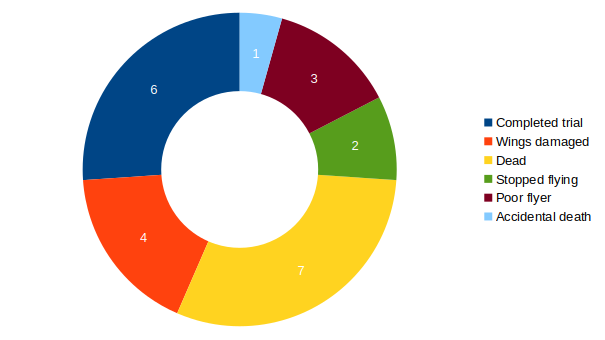
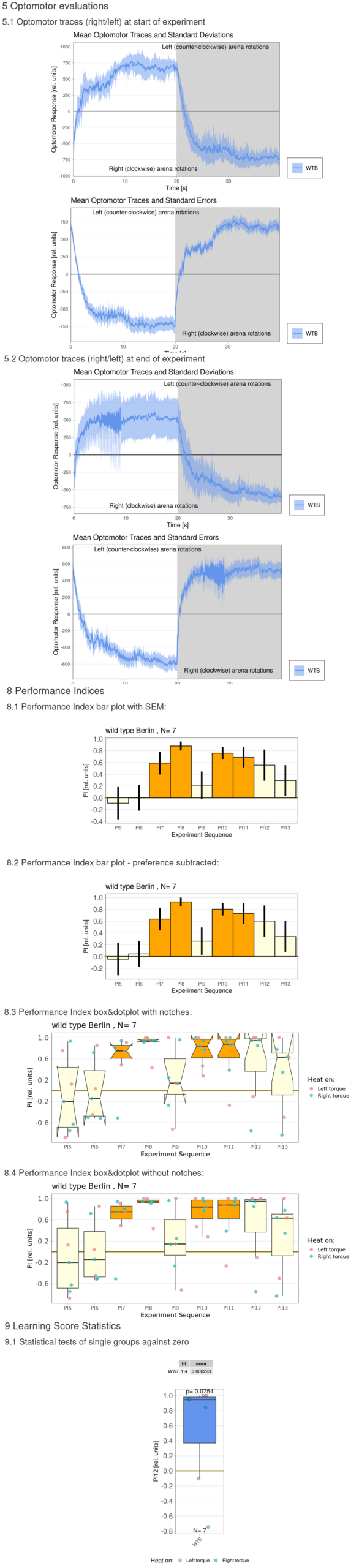
Torque measurements 10.02.2026
on Friday, February 13th, 2026 12:31 | by Fridrik Kjartansson
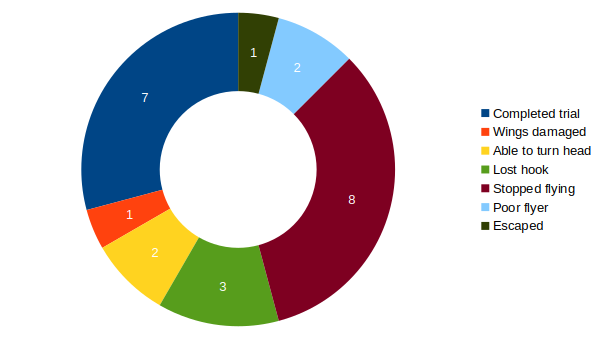
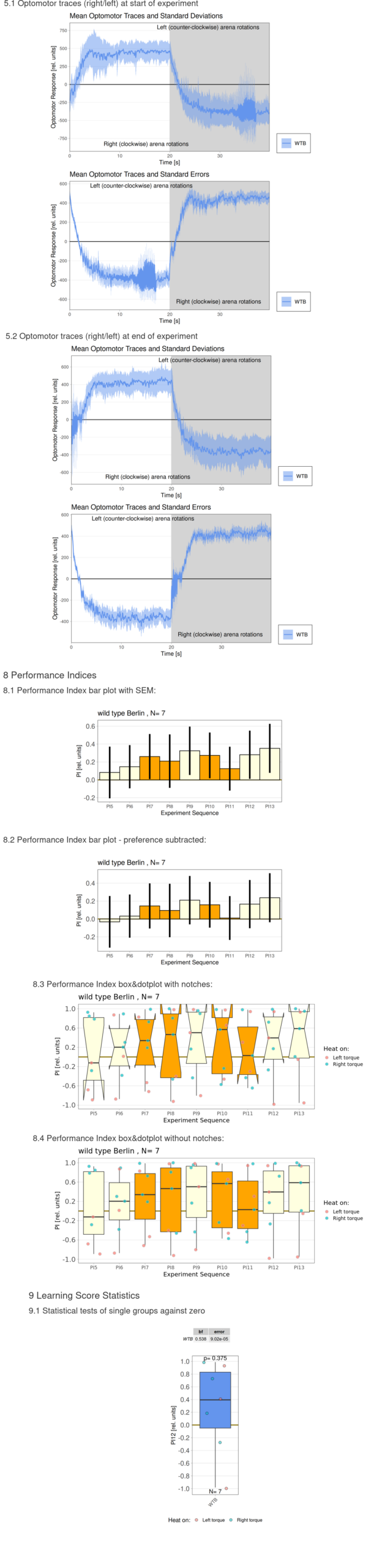
Torque measurements 2.2. – 6.2
on Monday, February 9th, 2026 12:41 | by Fridrik Kjartansson
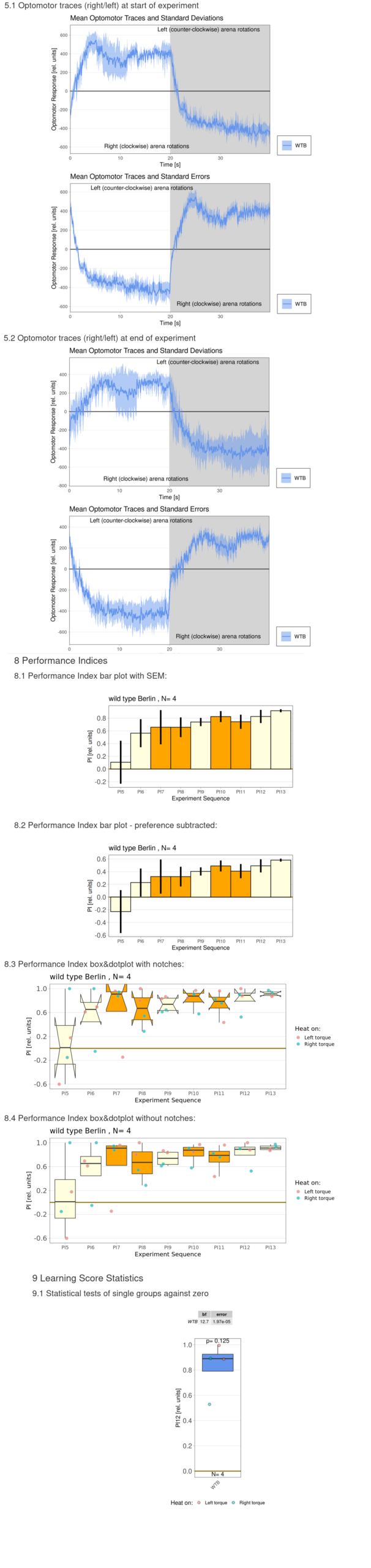
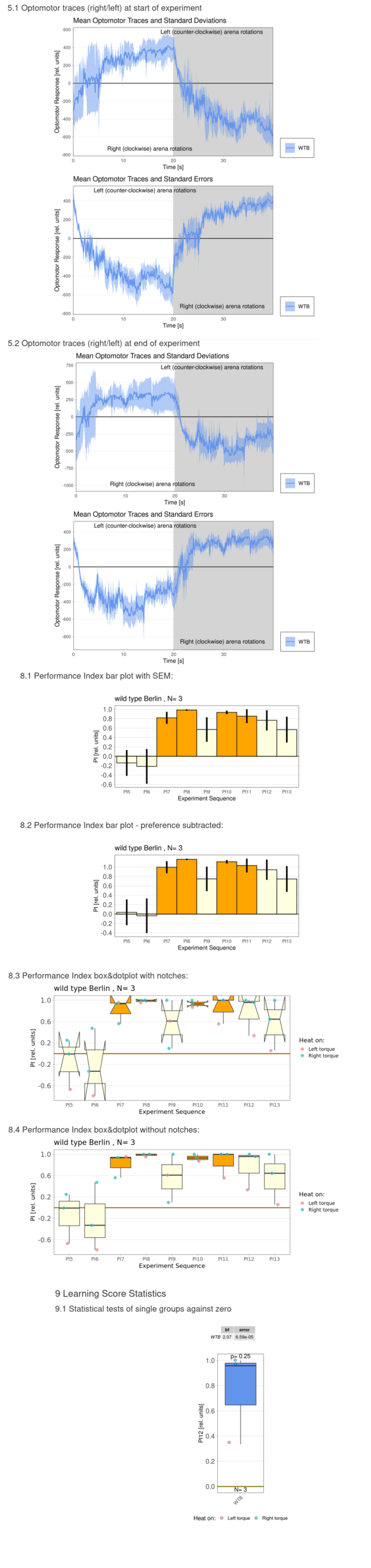
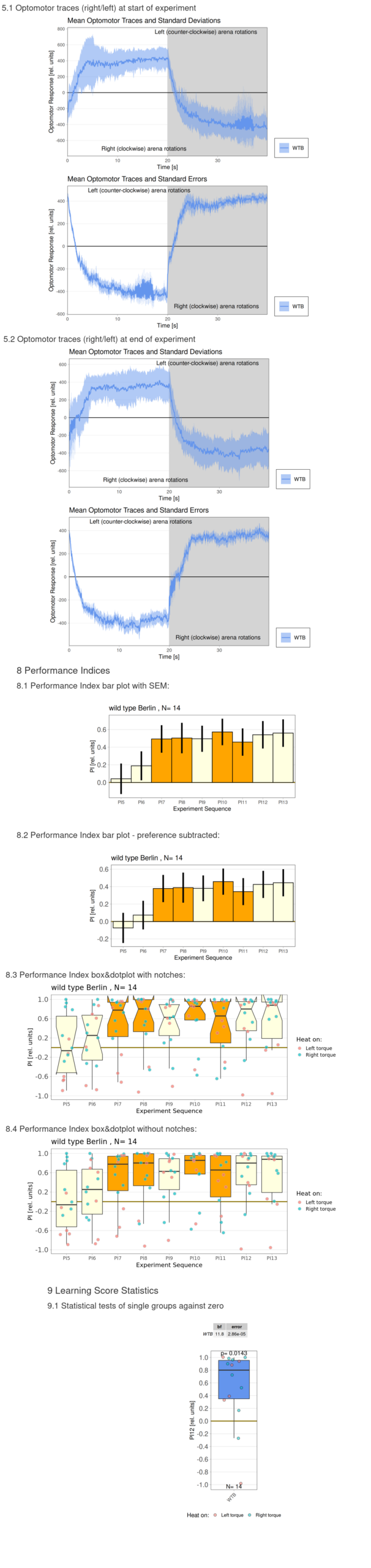
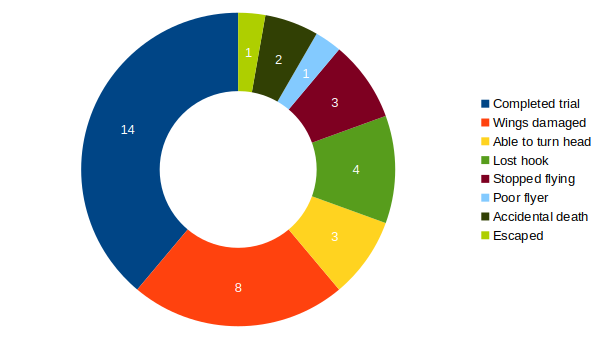
Category: flight, Habit formation, operant self-learning, Optomotor response | No Comments
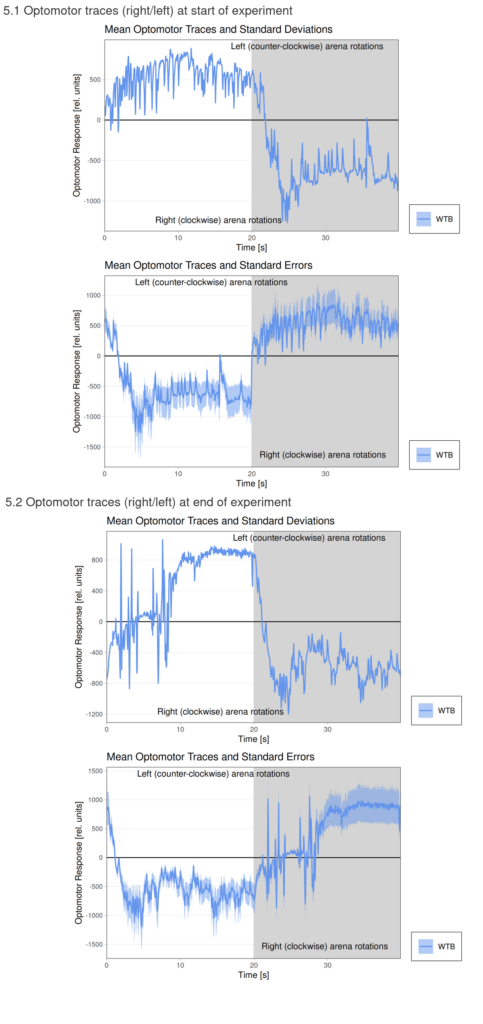
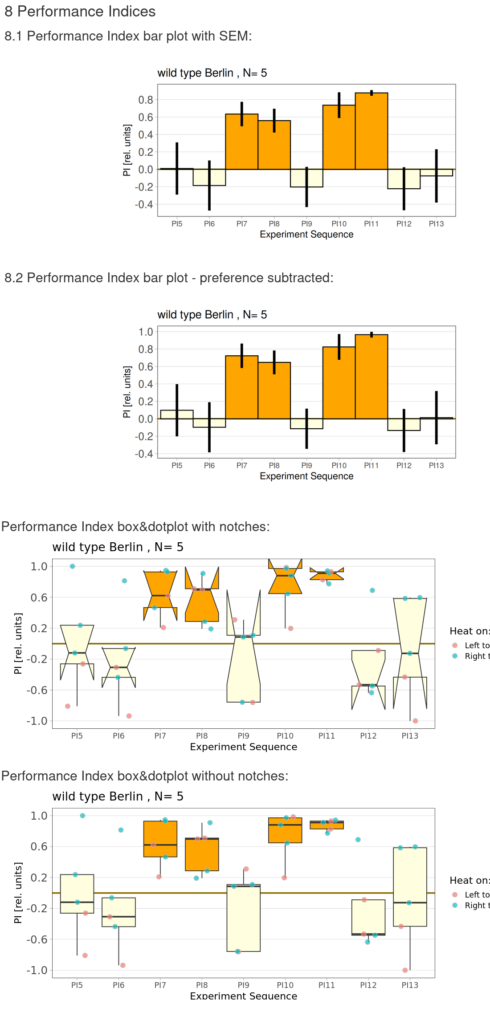
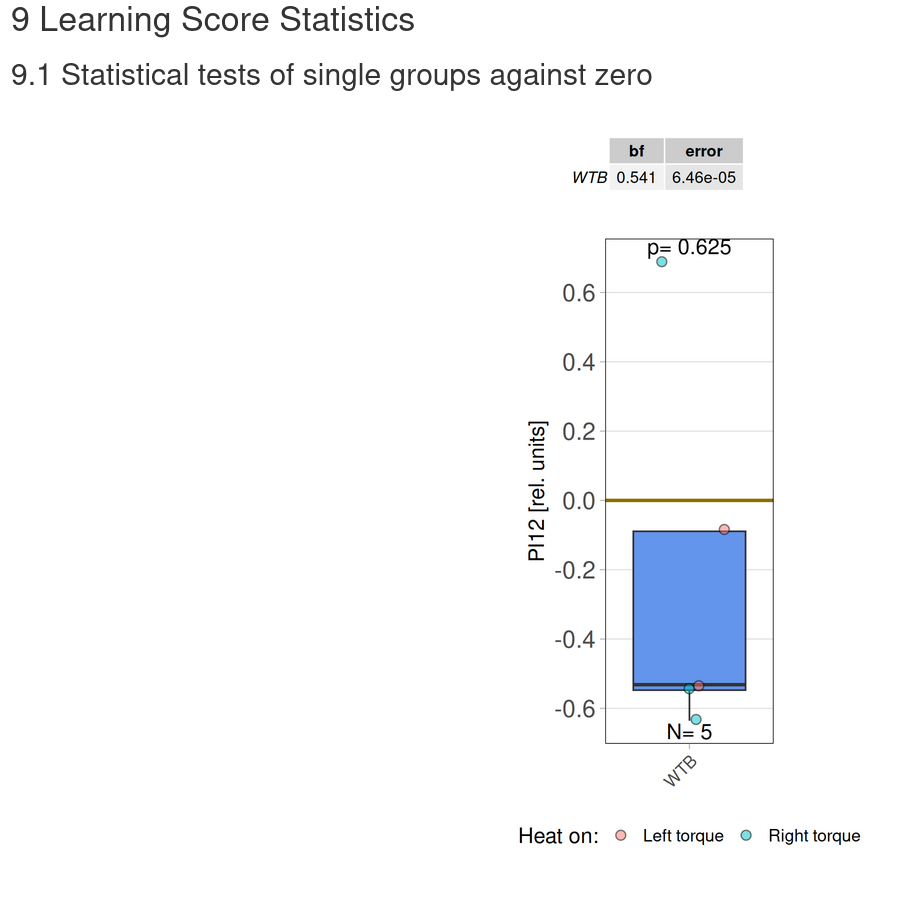
Torque measurements
on Monday, January 19th, 2026 10:32 | by Fridrik Kjartansson
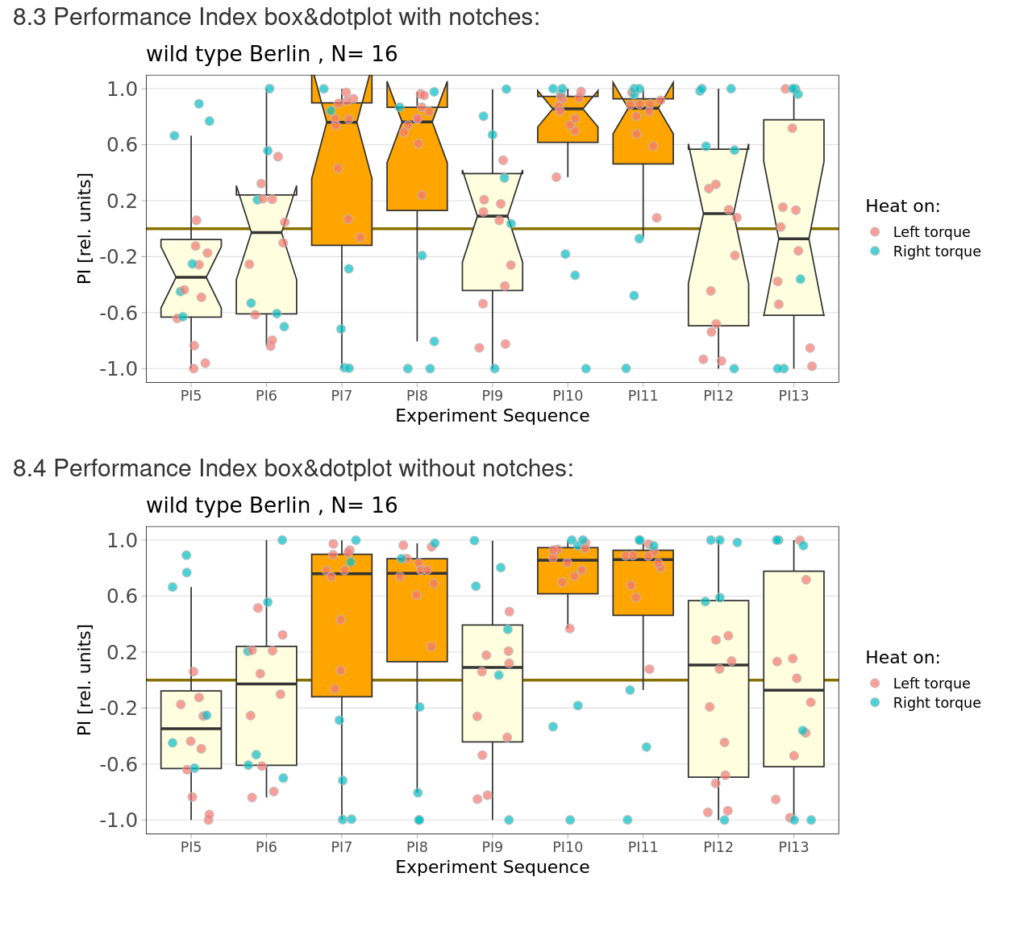
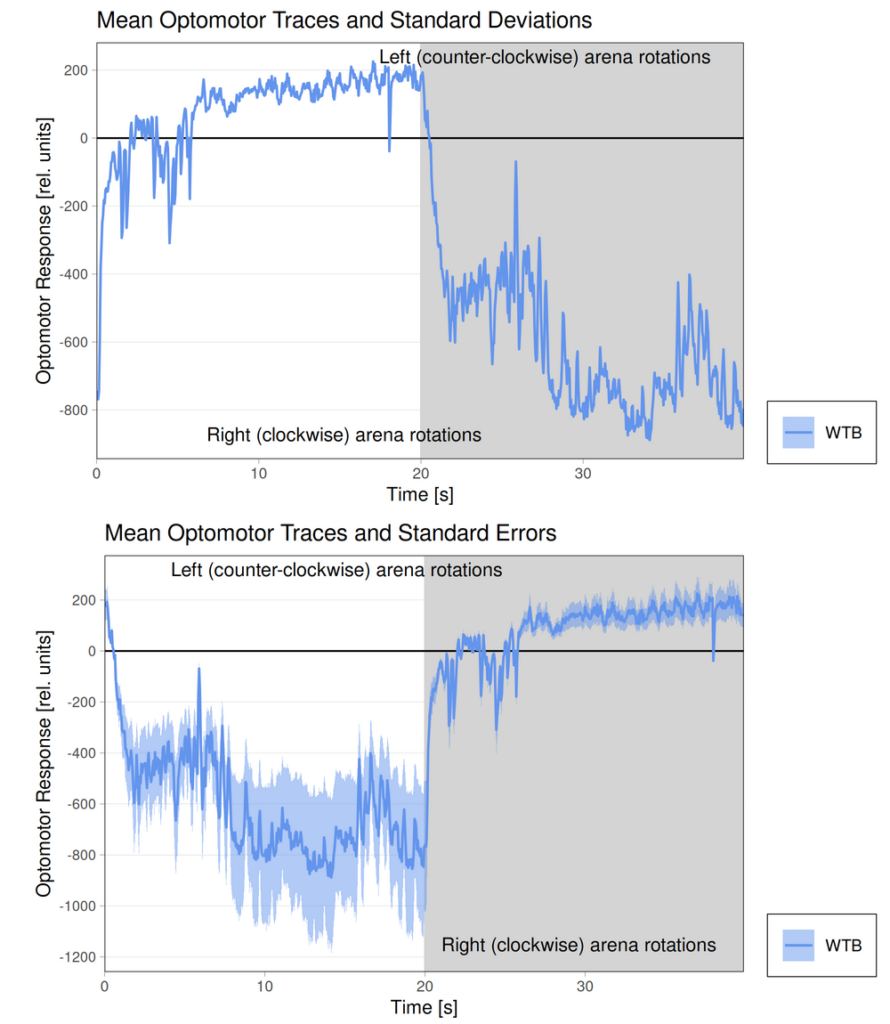
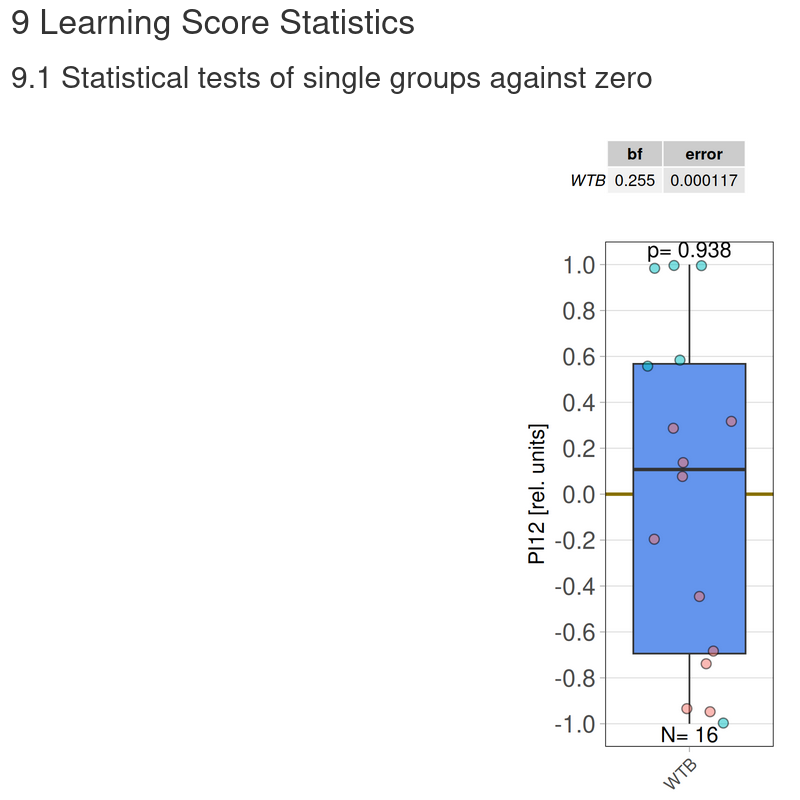
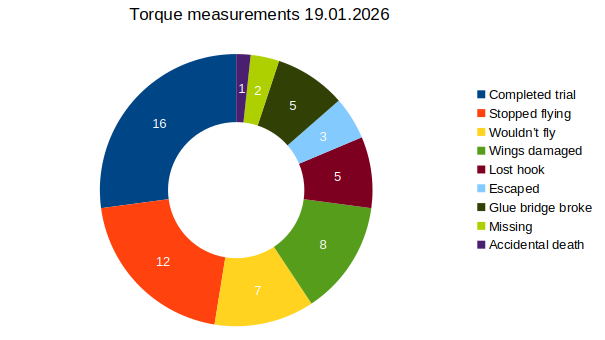
Category: flight, Memory, Operant learning, operant self-learning, Optomotor response | No Comments
Torque learning results 12.12.2025 FBK
on Monday, December 15th, 2025 10:52 | by Fridrik Kjartansson
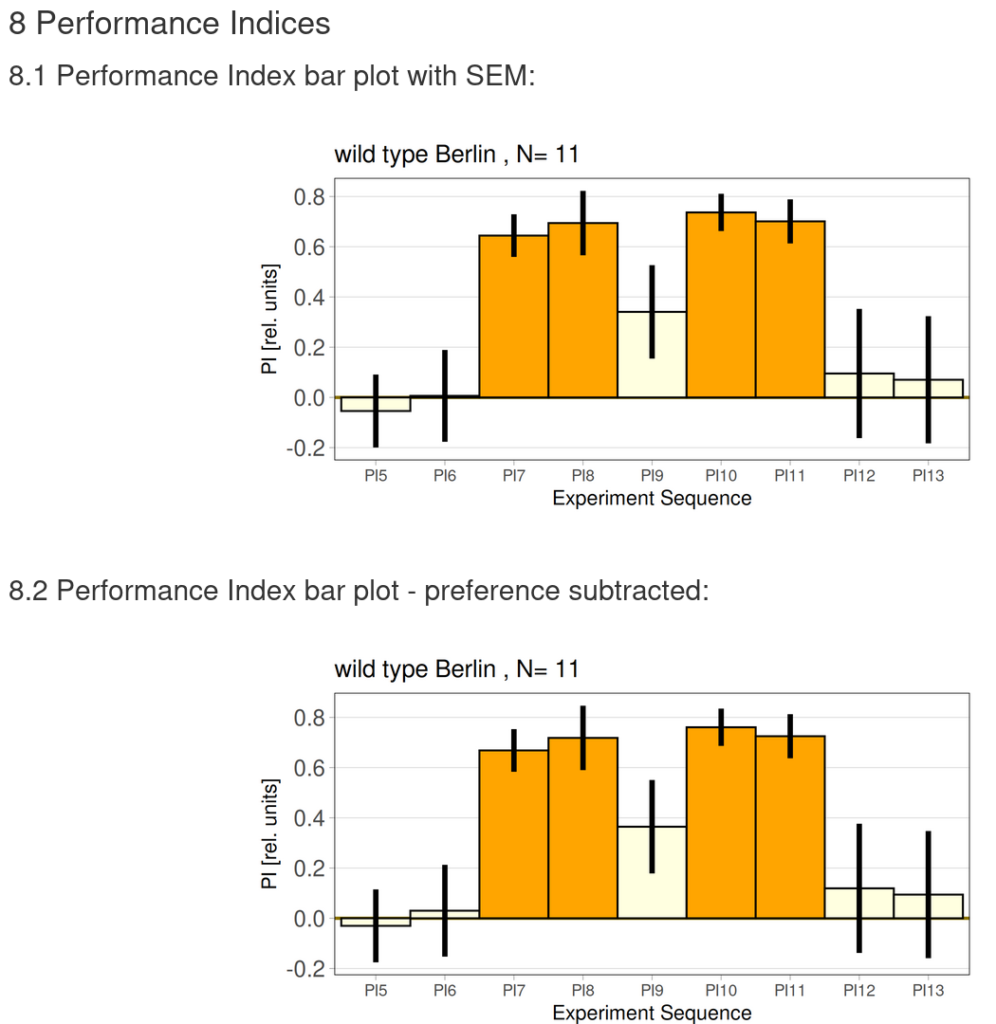
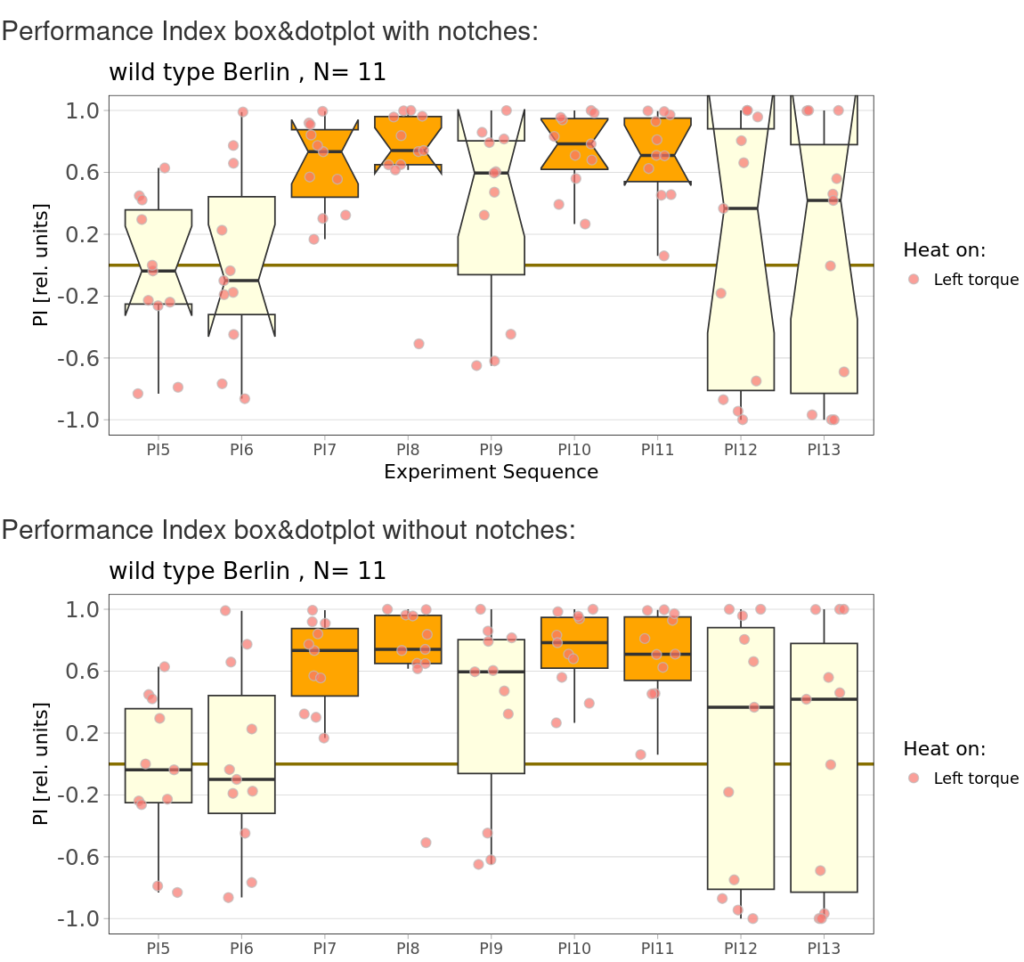
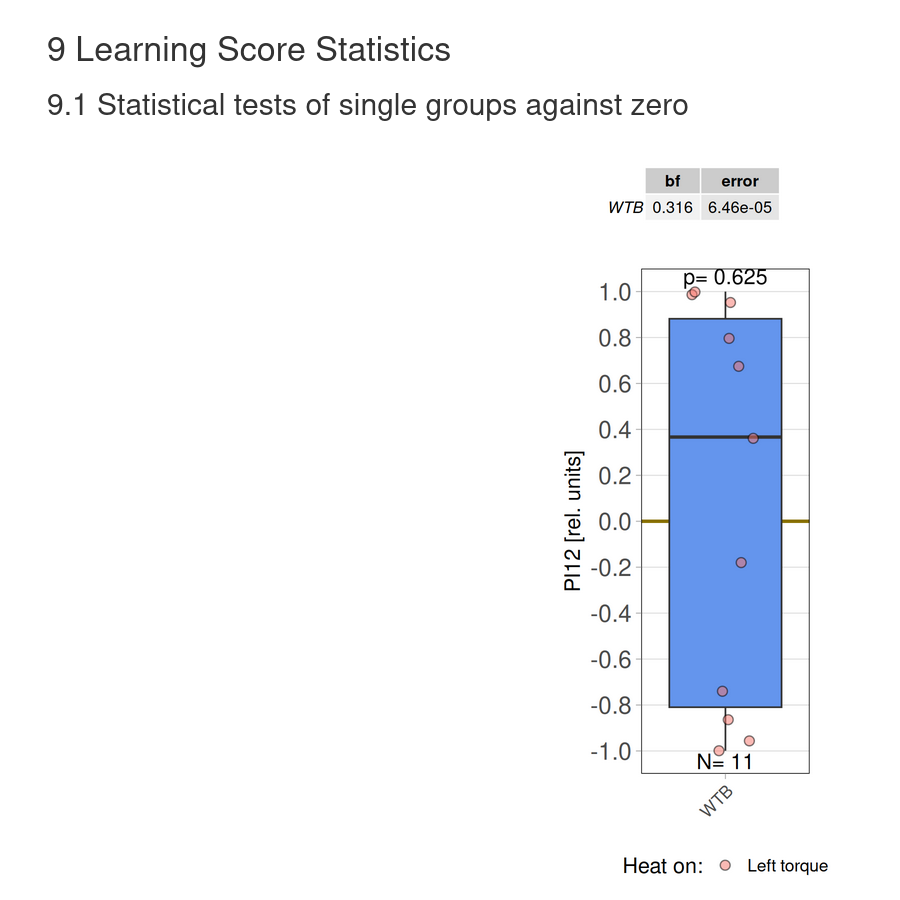
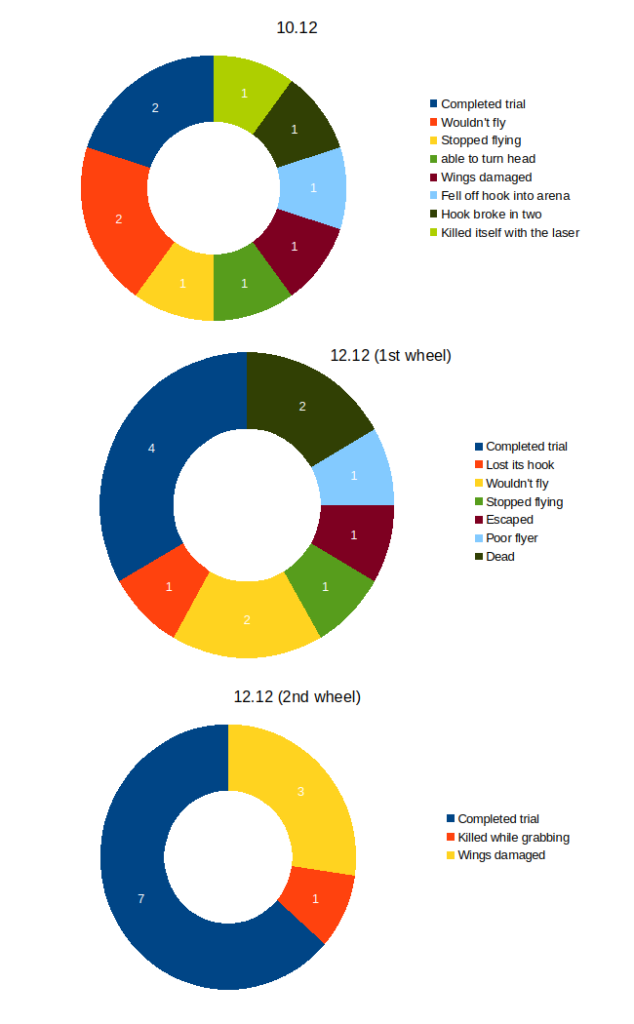
Category: flight, Operant learning, operant self-learning, Optomotor response | No Comments
Yaw torque learning
on Monday, August 4th, 2025 1:57 | by Julia Schulz
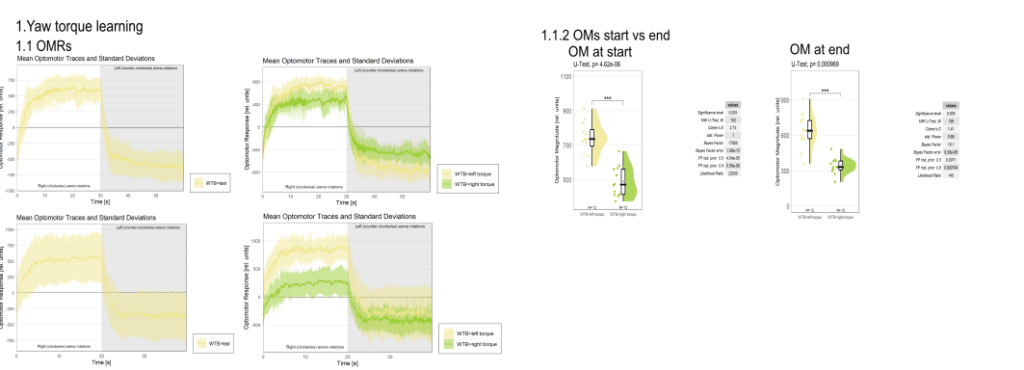
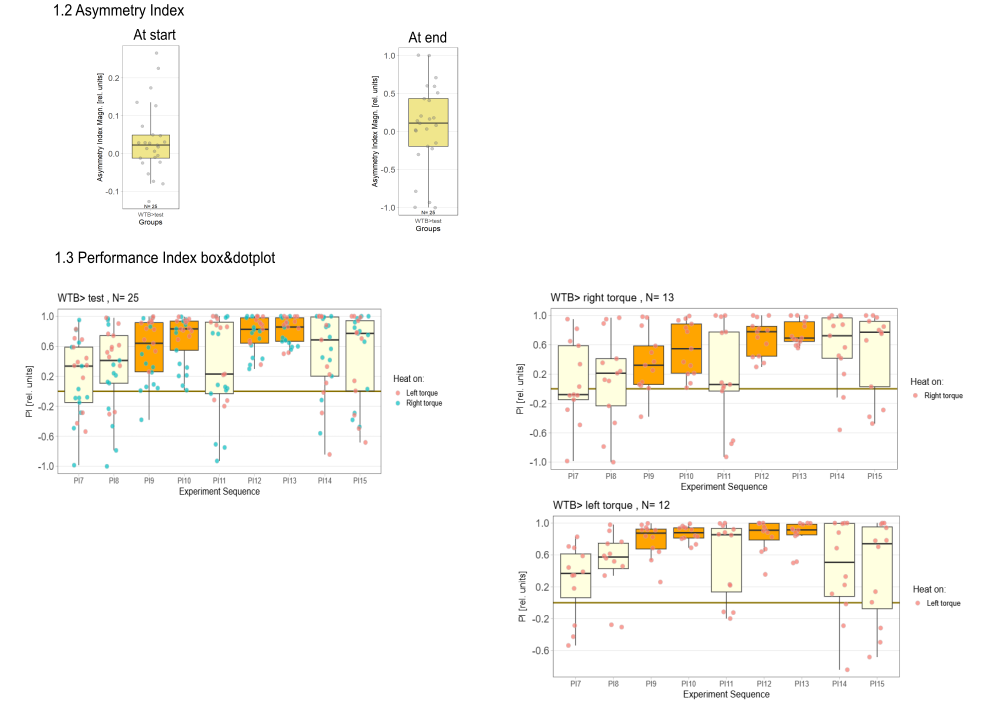
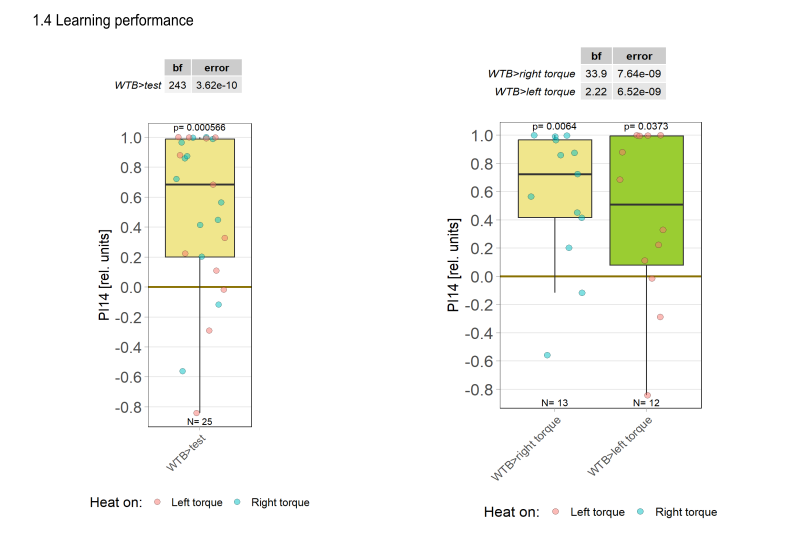
Category: Operant learning, operant self-learning, Optomotor response | No Comments