Adding results for T-Maze (and correcting y-axis description)
on Sunday, July 21st, 2024 5:15 | by Daniel Döringer
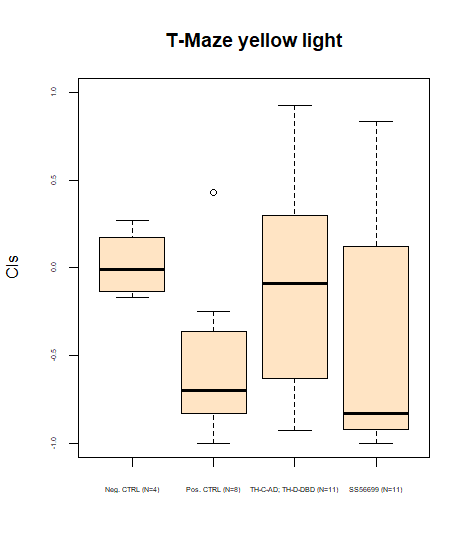

Category: Uncategorized | No Comments
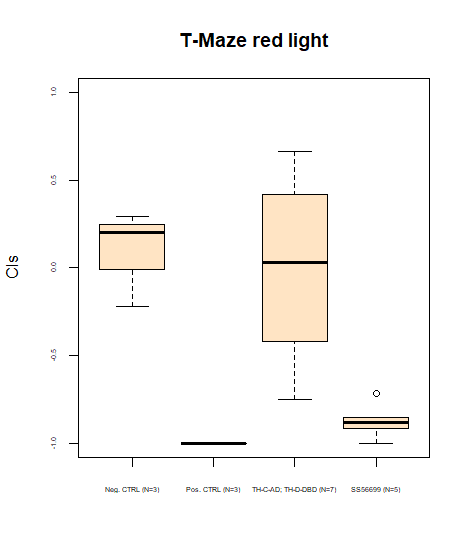
First T-Maze results coming in
on Monday, July 15th, 2024 8:41 | by Daniel Döringer

Category: Uncategorized | No Comments
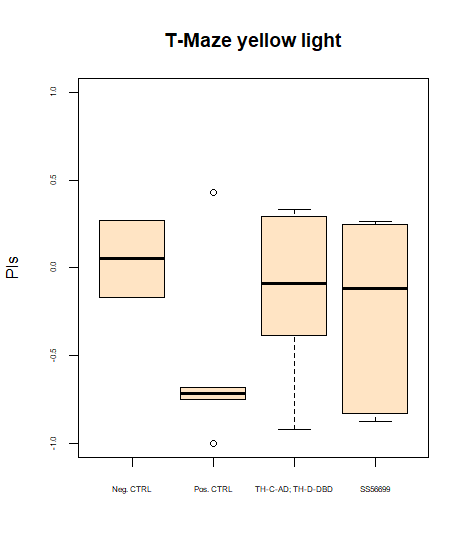
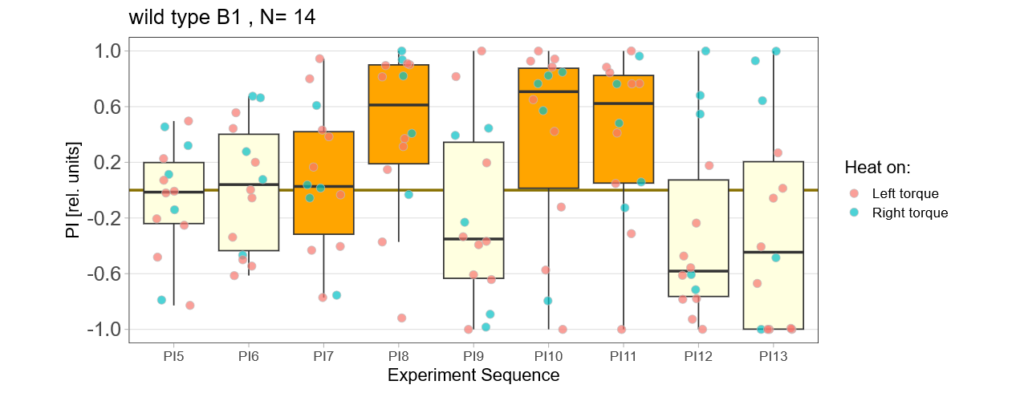
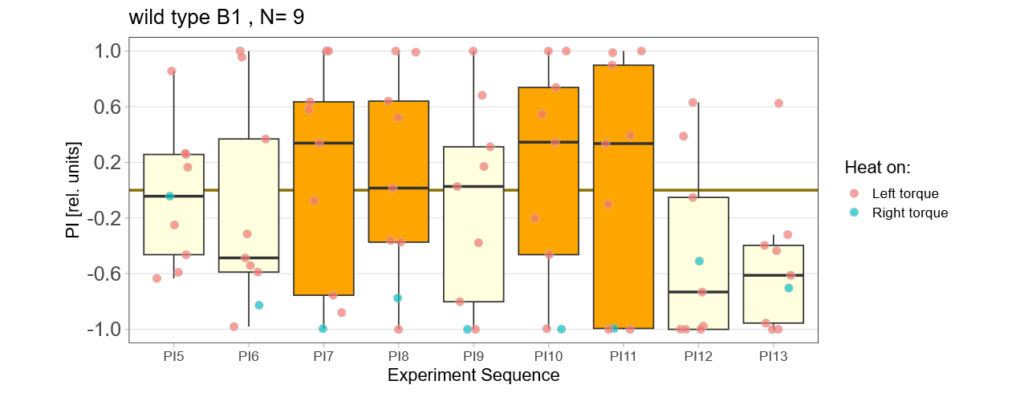
More data from the basement
on Monday, July 8th, 2024 8:15 | by Ellie
With the modified laser settings I was able to test 3 more wtb. Here are the results:

–> yaw torque
Results for Dop1R1-RNAi, Amon-RNAi and the course data
on Tuesday, May 21st, 2024 8:11 | by Maja Achatz
Firstly, I did an experiment where Dop1R1 was knocked down via RNAi. The following shows the results:

For the next experiment, the controls groups were the same, however in the experimental group Amon was down regulated. These are the results:

Lastly, in the course the data for the larvae experiment was collected only under blue light, so I used the same crossings and tested them under red light to use as a control.

Category: Kenyon cells, Larve, Mushroom Body, Optogenetics, Uncategorized | No Comments
First data from aPKC knock-out in FoxP neurons
on Monday, May 13th, 2024 8:49 | by Björn Brembs
When Andi tested his aPKC CRISPR knock-outs, he used the Tang torque meter setup, where the OMRs aren’t recorded. So I’m attempting to replicate his results and compare OMRs between groups.

It looks like I need to tweak the graph in some way to make the density plots show up. Obviously, N is too low to say anything yet, but I need to work on the plot.
Category: Uncategorized | No Comments
Bachelor Blog / #5 no learning :(
on Monday, September 11th, 2023 12:21 | by Ellie
Below you find the data from my experimental rounds A and B:
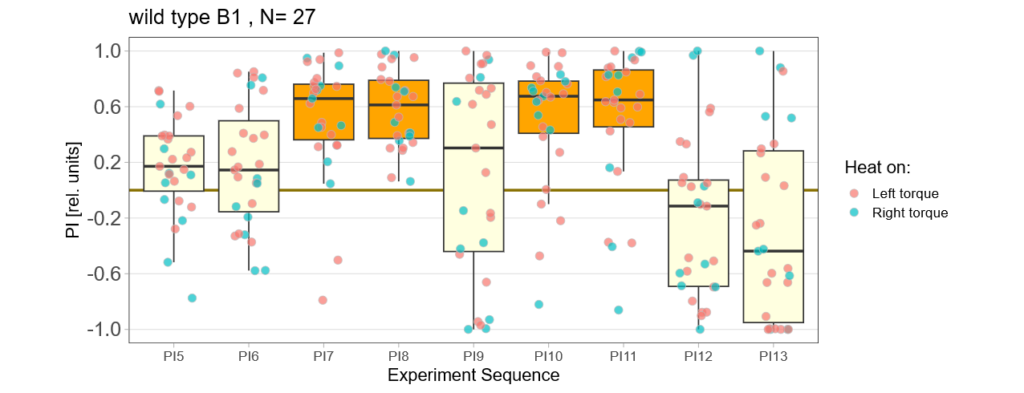
-> learning scores of the parental flies from experimental round A and B
-> learning scores of the trained parent´s offspring only from experimental round A
-> learning scores of the untrained parent´s offspring only from experimental round A
The results confuse me a lot and I am happy to discuss reasons :) However the offspring of the round-B will be ready for testing by the end of this week so there is still some data to collect…
Success: rsh Stock has rsh1 Mutation
on Monday, August 7th, 2023 11:11 | by Isabel Stark
Via gDNA analysis and PCR was the specific area of the rsh gene extracted and amplified where the nucleotide substitution: C to T (Folkers et al., 2006) should be for the rsh1 mutation. The amplicon was Sanger sequenced which proved the nucleotide substitution.
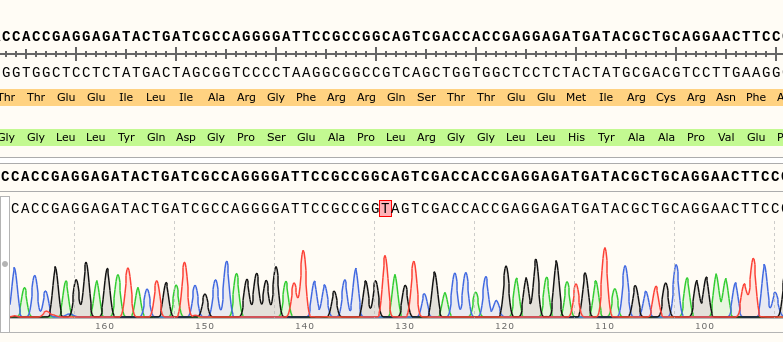
Category: genetics, Memory, Operant learning, operant self-learning, Radish, Uncategorized | No Comments
Creating gRNAs via PCR
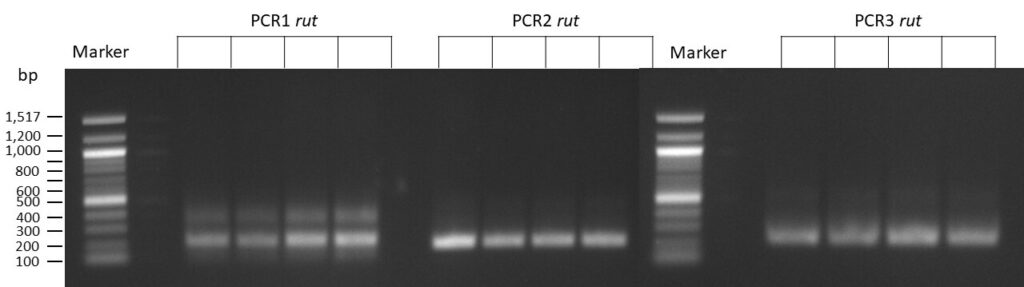
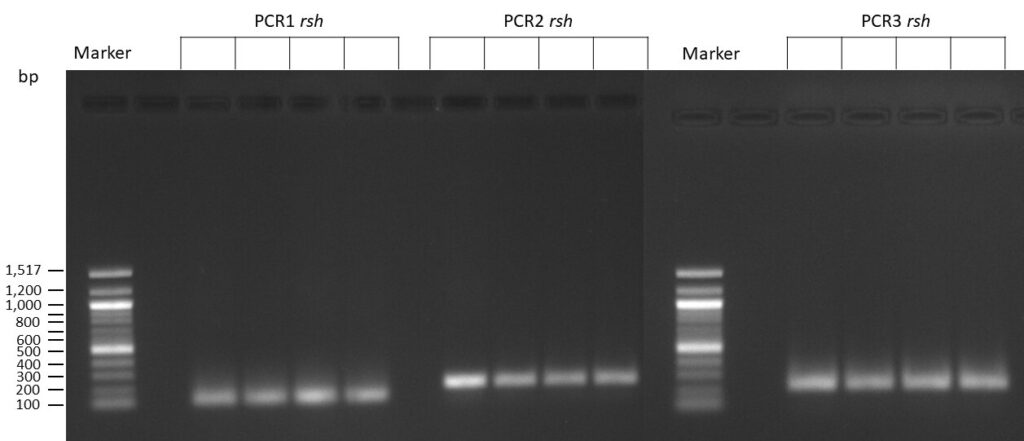
on Friday, July 7th, 2023 6:38 | by Isabel Stark
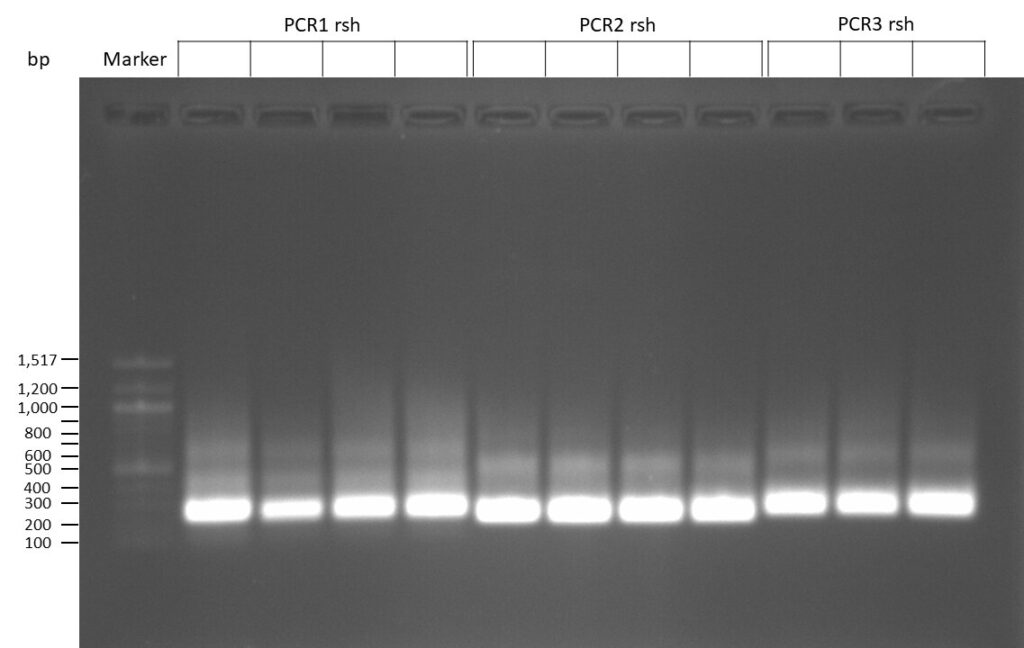
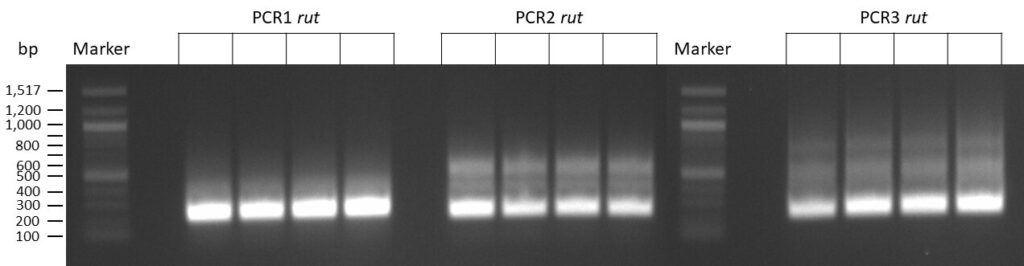
The template pCFD6 was used with a concentration of 640 pg/µl.
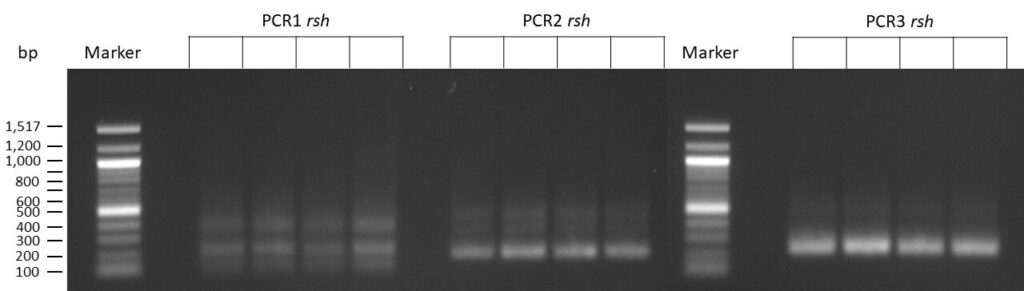
The template pCFD6 was used with a concentration of 64 pg/µl (1:10 dilution).
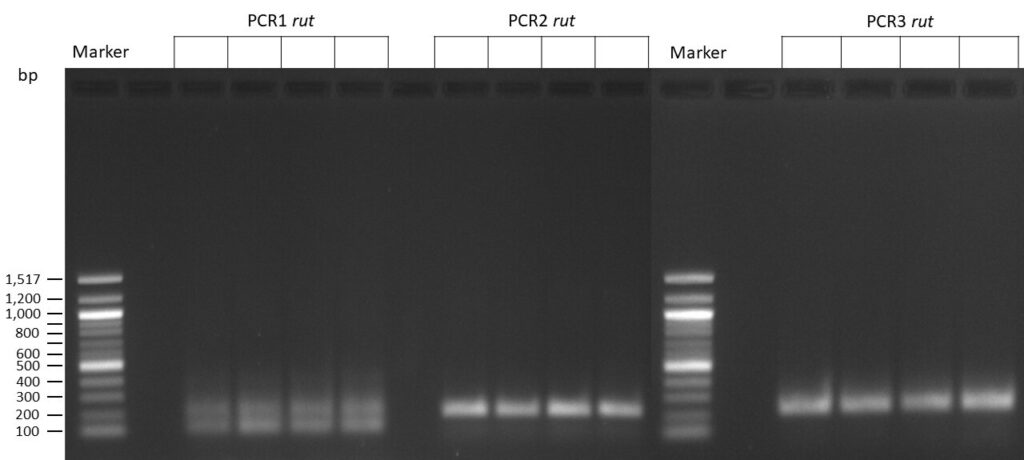
The template pCFD6 was used with a concentration of 64 pg/µl (1:10 dilution).
50µl of 5xQ5 High GC Enhancer was added to the PCR mix.
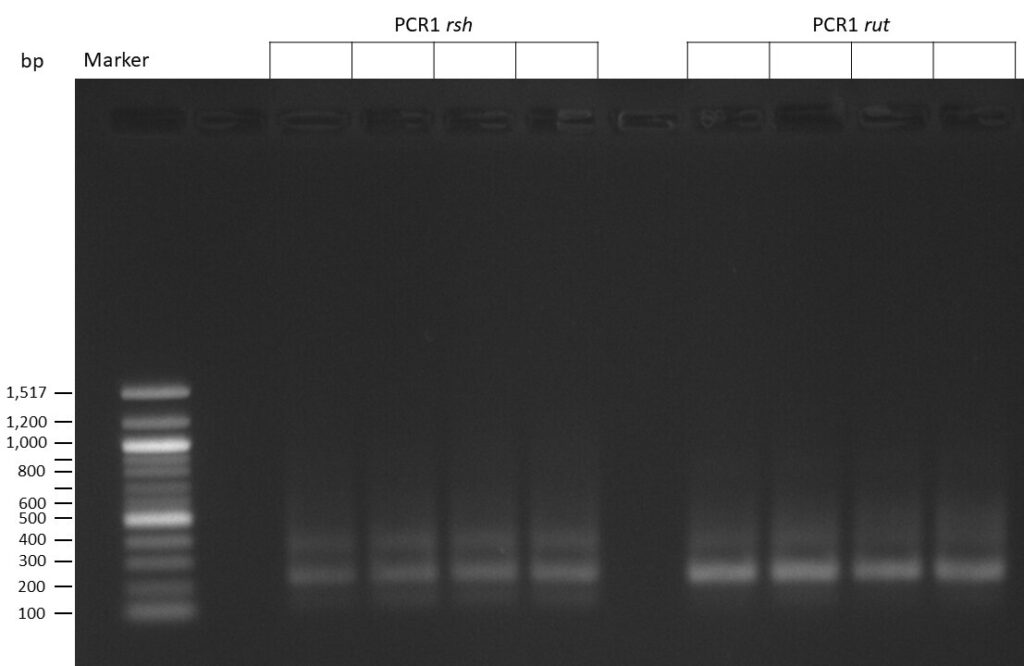
The template pCFD6 was used with a concentration of 128 pg/µl (1:5 dilution).
50µl of 5xQ5 High GC Enhancer was added to the PCR mix.
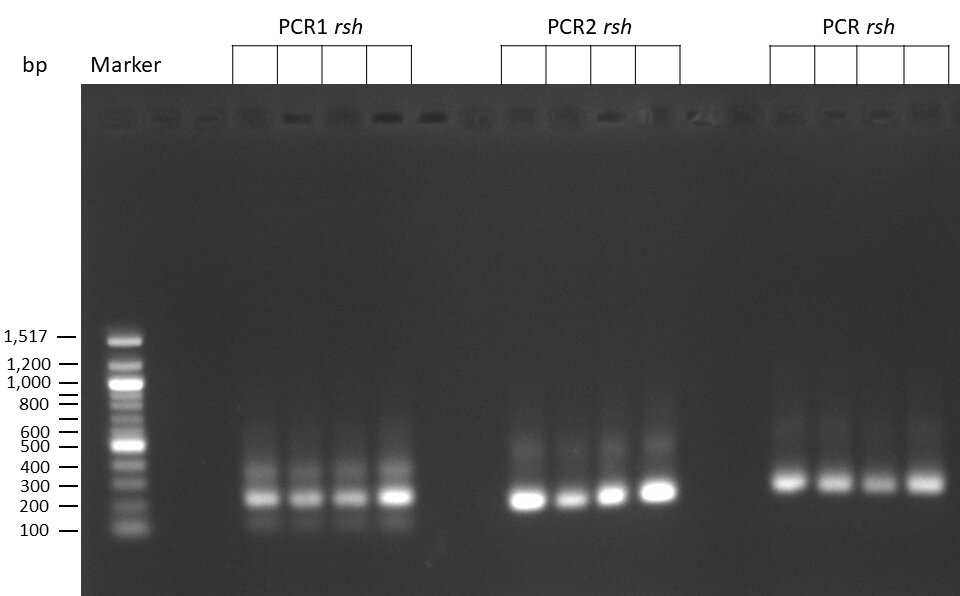
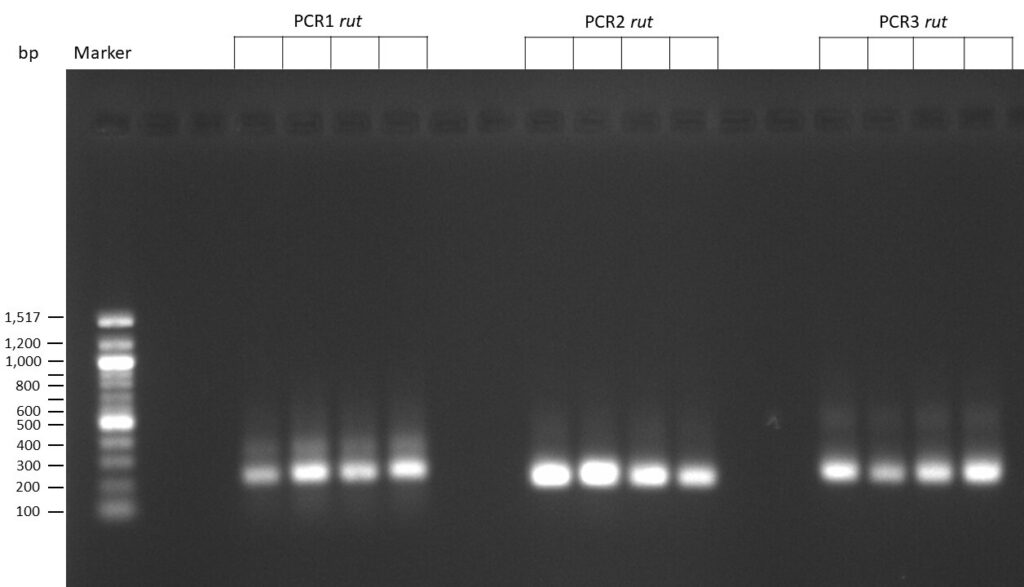
The template pCFD6 was used with a concentration of 640 pg/µl.
The Phusion DNA Polymerase was used instead of the Q5 High-Fidelity DNA Polymerase.
Category: genetics, Memory, Operant learning, operant self-learning, Radish, Uncategorized | No Comments
Flight Duration
on Monday, January 30th, 2023 12:40 | by Silvia Marcato

TntG expression
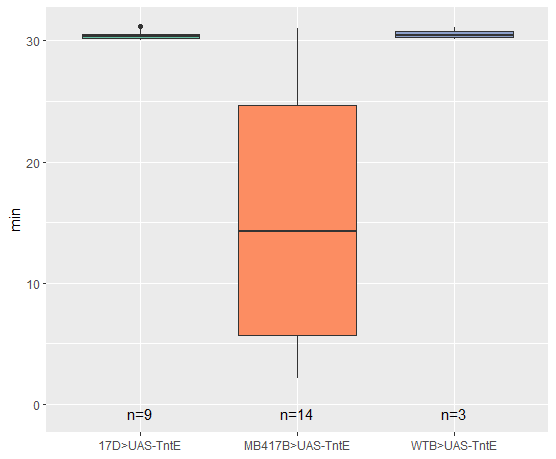
TntE expression
Category: Uncategorized | No Comments
Kibra knockout, adult
on Monday, December 19th, 2022 11:15 | by Andreas Ehweiner
Knockout of Kibra in adult flies.
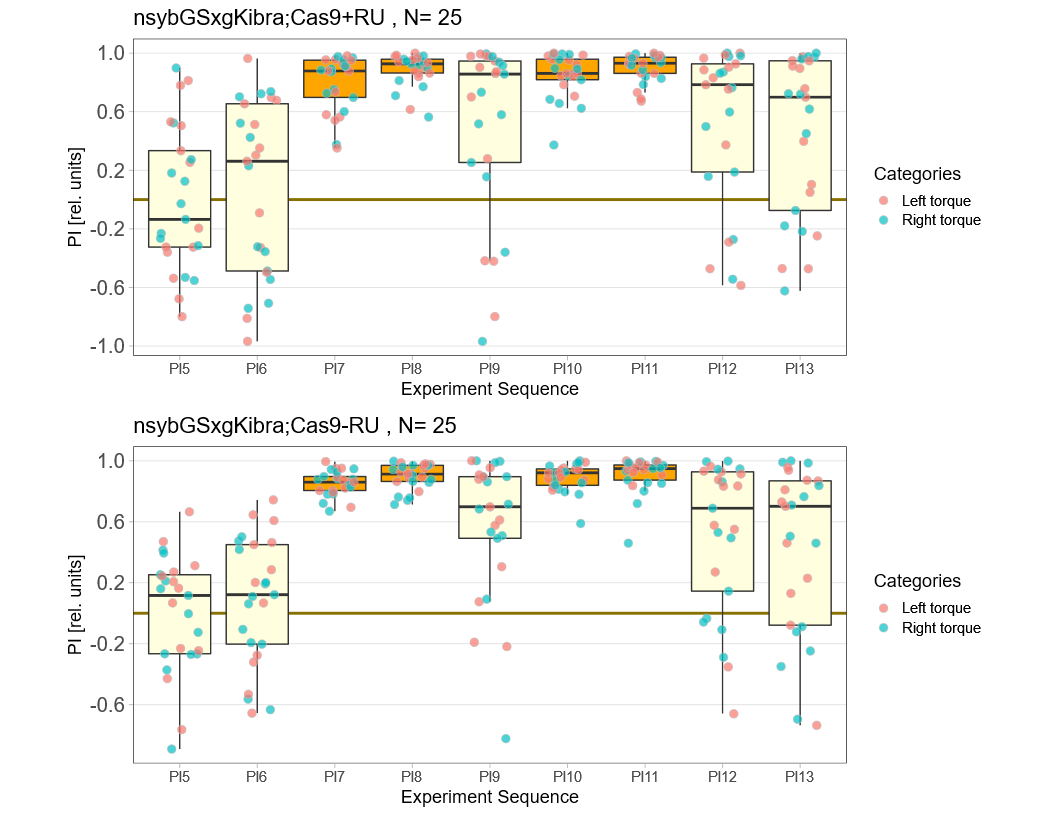
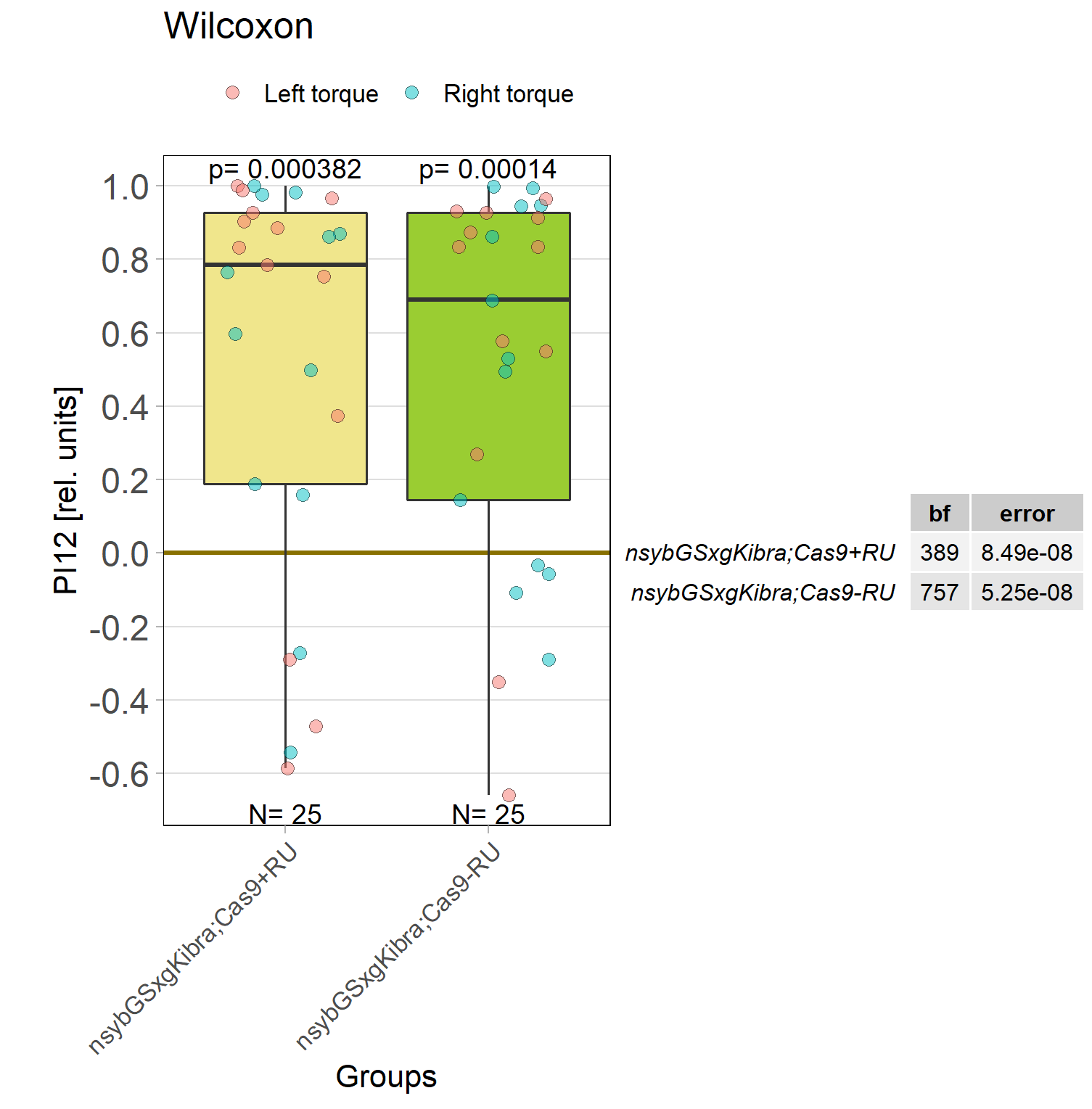
Category: flight, Memory, Operant learning, operant self-learning, Uncategorized | No Comments