Best intensity of light for crimson channel in Joystick experiment
on Wednesday, June 13th, 2018 12:27 | by Saurabh Bedi
Used positive control flies (NorpA; UAS Crimson) to check what intensity of light is better for activating the crimson channel and for studying learning behaviour in the joystick experiment. Tested light in 3 intensity ranges to check what would be best suited.
- Initially, we were doing experiments at 700 lux intensity. We saw results which possibly point out to damaging of neuronal populations due to such high-intensity light. As can be seen in the graph below, there is a strong aversion in the first 2 minutes of the training period. However, this aversion drastically decreases later and a very strong phenotype is not shown later. This may prove the hypothesis that high-intensity
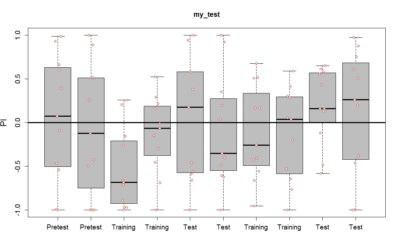 light(about 700 lux) may result in damage to neuronal populations.
light(about 700 lux) may result in damage to neuronal populations. - So we tried with two more light intensities – 400 lux and 100 lux. The graphs for these are att
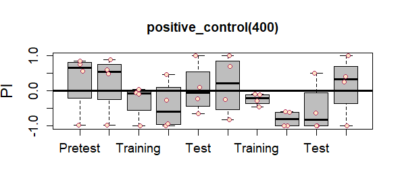
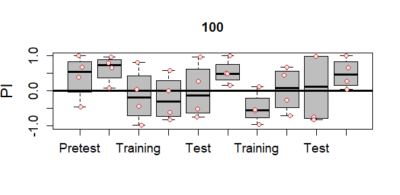 ached below. As can be clearly seen that now the results dont just show up for the first training segment but remain til the very end. This indicates that both 100lux and 400 lux light intensity are safe and do not disrupt neurons. Also, it can be clearly seen that there is a stronger and more pronounced effect with 400 lux light intensity. Thus we decide that 400 lux light intensity needs to be used for testing crimson channel expression for the joystick experiment.
ached below. As can be clearly seen that now the results dont just show up for the first training segment but remain til the very end. This indicates that both 100lux and 400 lux light intensity are safe and do not disrupt neurons. Also, it can be clearly seen that there is a stronger and more pronounced effect with 400 lux light intensity. Thus we decide that 400 lux light intensity needs to be used for testing crimson channel expression for the joystick experiment.

Category: neuronal activation, Optogenetics | No Comments
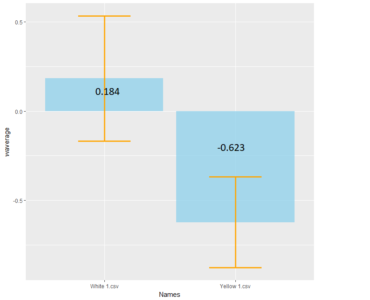
Tmaze experiments : Test for Blindness and Comparison with positive control
on Monday, June 11th, 2018 12:45 | by Naman Agrawal
Comparison between White 1 ( Control (NorpA- UAS Chrimson)) and Yellow 1 (Positive Control ( Gr28bd-G4, TrpA1-G4).
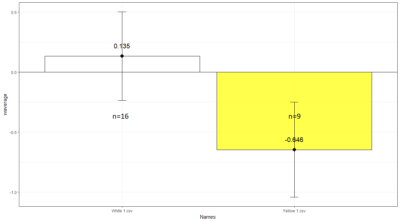
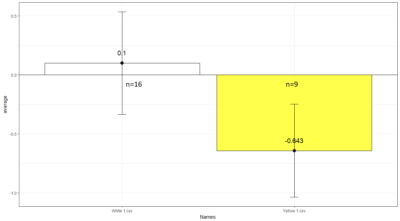
To test whether the flies are really blind, and there is no problem with the NorpA part of the construct, we compare with NorpA,UAS GTACR1 ; NorpA,UAS GTACR2 ; and another stock of NorpA-UAS-Chrimson.
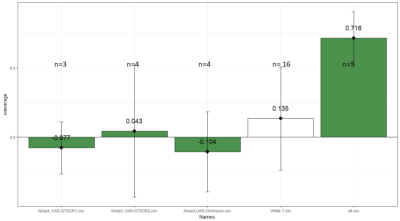
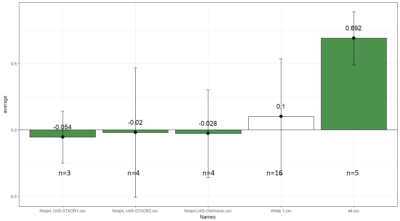
Weighted mean is calculated by multiplying the weights (total number of flies in that experiment/total number of flies in all the experiments) with the PI for that experiment, and taking the sum thereafter for all the experiments.
Category: genetics, neuronal activation, Optogenetics, TrpA | No Comments
Tmaze experiments initial results
on Monday, June 4th, 2018 2:16 | by Naman Agrawal
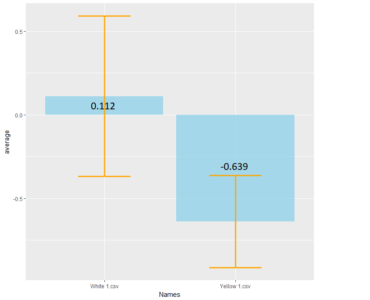
 the first graph shows Bar plot of Mean and standard deviation of PIs for Genetic control (n=9) and Positive control (n=5).
the first graph shows Bar plot of Mean and standard deviation of PIs for Genetic control (n=9) and Positive control (n=5).
The second graph shows weighted mean and weighted standard deviation of the same.
Weighted mean is calculated by multiplying the weights (total number of flies in that experiment/total number of flies in all the experiments) with the PI for that experiment, and taking the sum thereafter for all the experiments.
Category: neuronal activation, operant self-learning, Optogenetics, TrpA | No Comments
Thornlabs spectrometer working!
on Monday, November 20th, 2017 2:59 | by Christian Rohrsen
After getting a new spectrometer, we confirmed that the first one was faulty. Comparison of old (1st and 3rd measures) and new (2nd and 4th) spectrometer. Now much more sensitive and the right spectrum measured for the green LED present in the spectrometer itself and for the light comming out of the light guide coupled with the red LED (whose spectrum does not seem to change after travelling through the light guide)
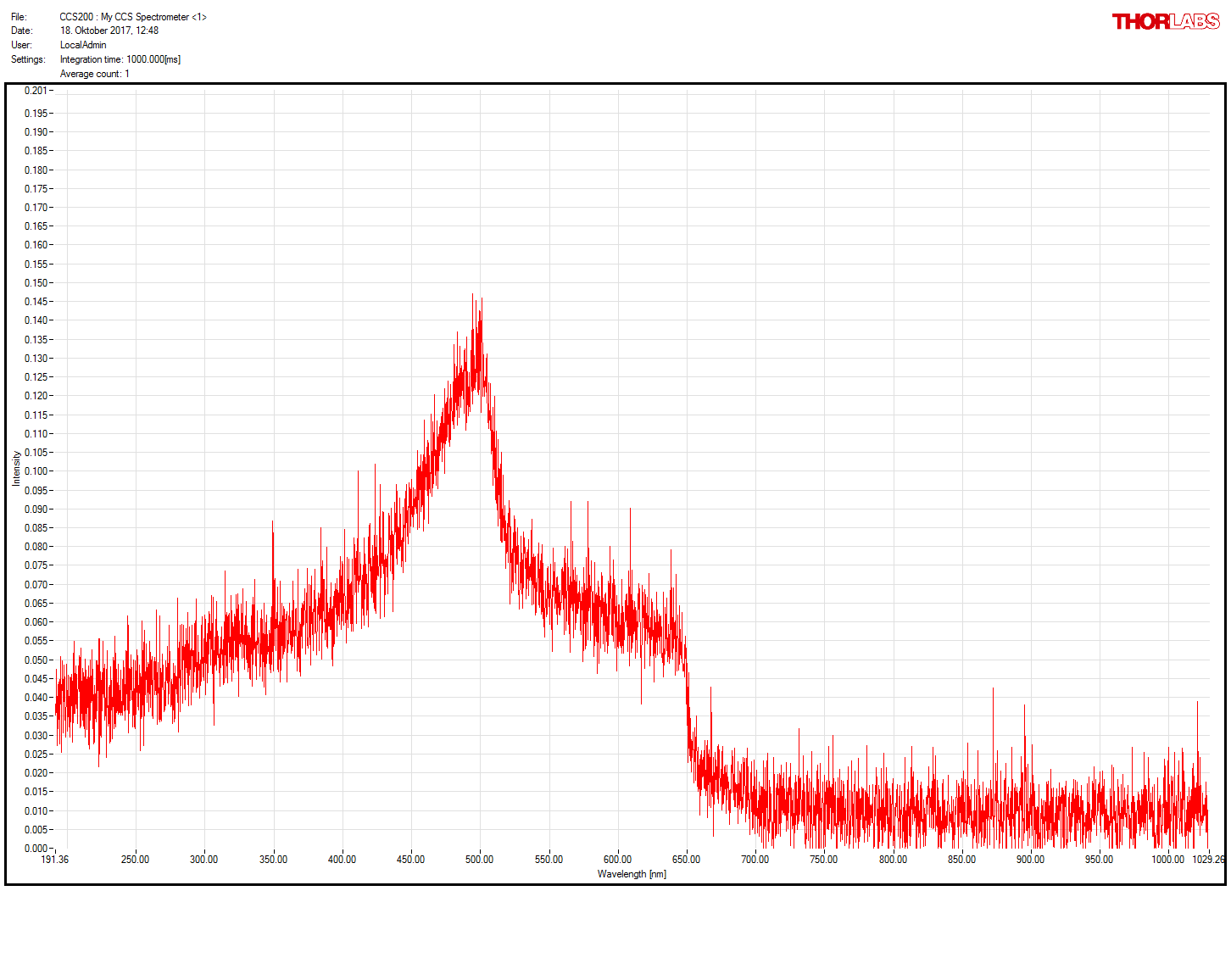
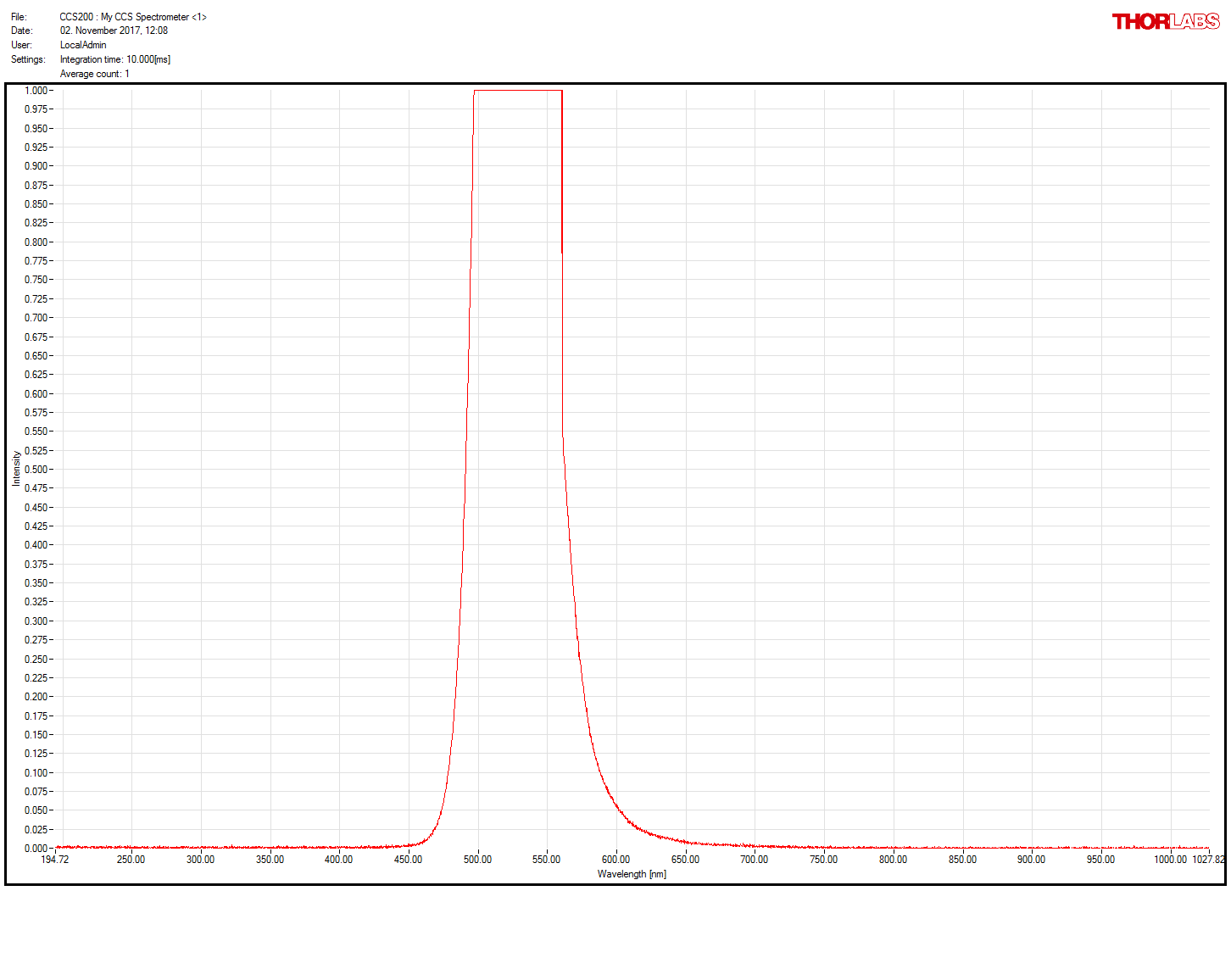
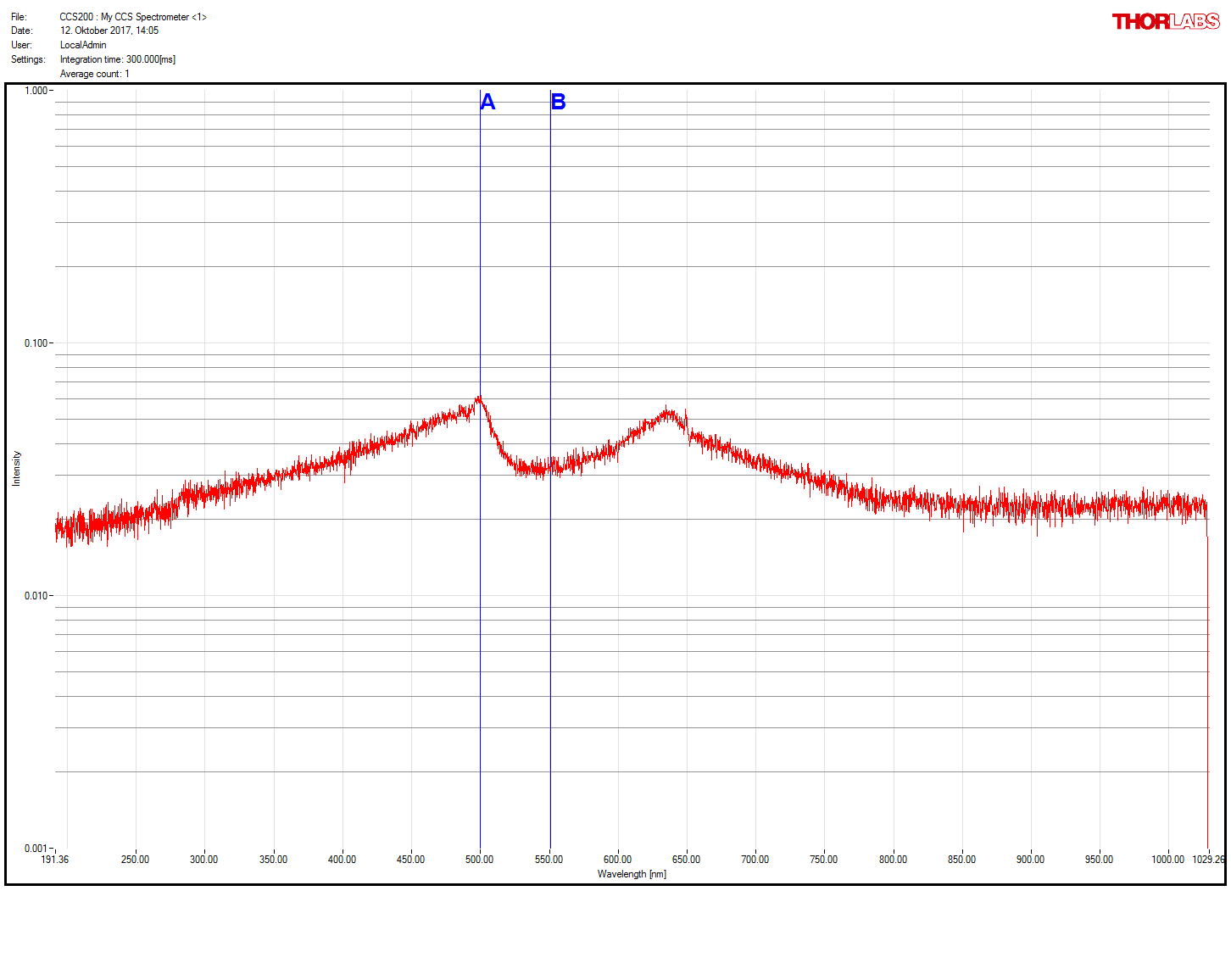

Category: neuronal activation, Operant reinforcment, Optogenetics | No Comments
thf1 line from screen
on Monday, July 17th, 2017 2:40 | by Christian Rohrsen
 This is the first line I completed from the screen. What it is more important is that I found out that NorpA;UAS-Chrimson has orange eyes, when it should be red. So I think I will start to do the line again and meanwhile test the contaminated line to see what the phenotype looks like
This is the first line I completed from the screen. What it is more important is that I found out that NorpA;UAS-Chrimson has orange eyes, when it should be red. So I think I will start to do the line again and meanwhile test the contaminated line to see what the phenotype looks like
Category: neuronal activation, Operant reinforcment, Optogenetics, python code | No Comments
Looking for students
on Monday, June 19th, 2017 6:01 | by Christian Rohrsen

We need a student to finish up experiments
Category: neuronal activation, Operant reinforcment, Optogenetics | No Comments
Measuring locomotion in the Joystick? Something seems to happen…
on Monday, June 19th, 2017 2:39 | by Christian Rohrsen
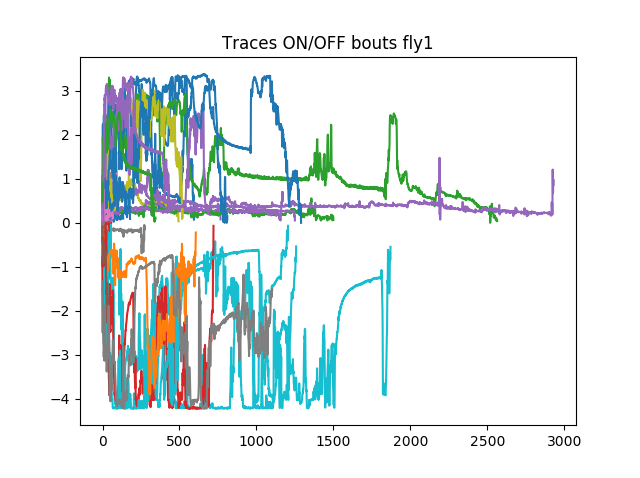
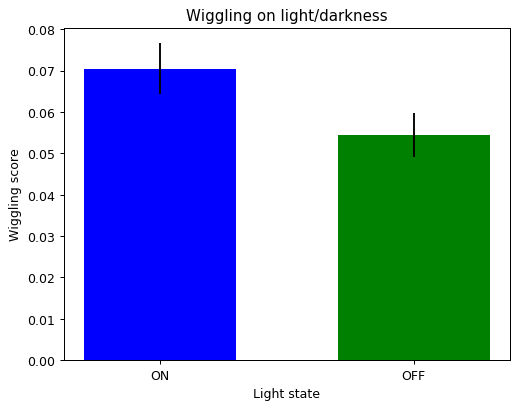
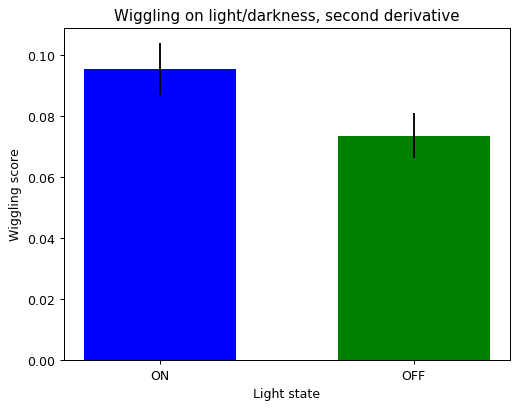
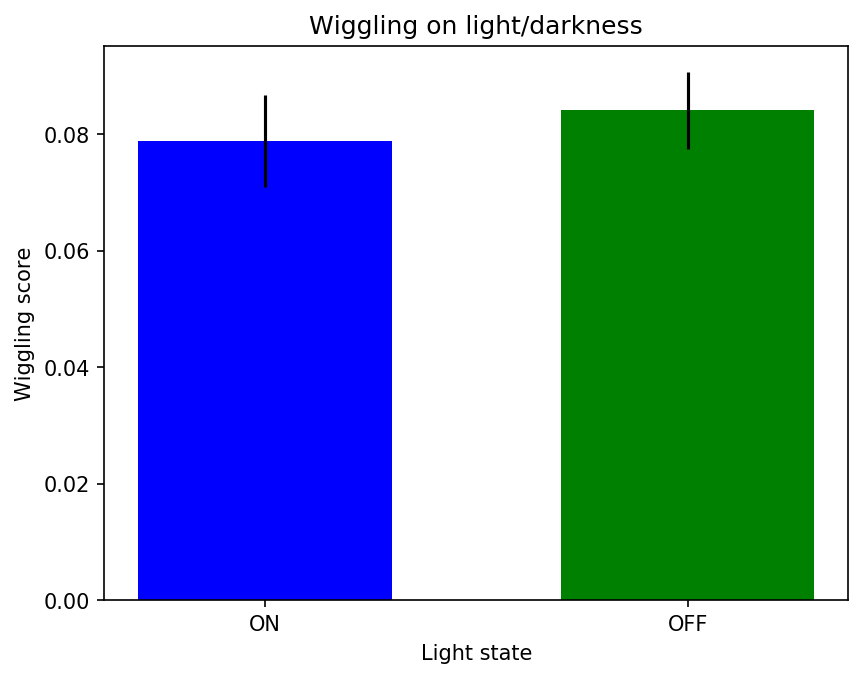
Example of ON/OFF traces. And then the wiggling was measured in both conditions (with and without light). It seems like they move more when they are in the light side. I also did measure the derivative with a tau=2 (in case the sampling is over the temporal frequency of fly behavior)
Category: neuronal activation, Operant reinforcment, Optogenetics, python code | No Comments
Comparing T- and Y-maze
on Monday, June 12th, 2017 2:59 | by Christian Rohrsen
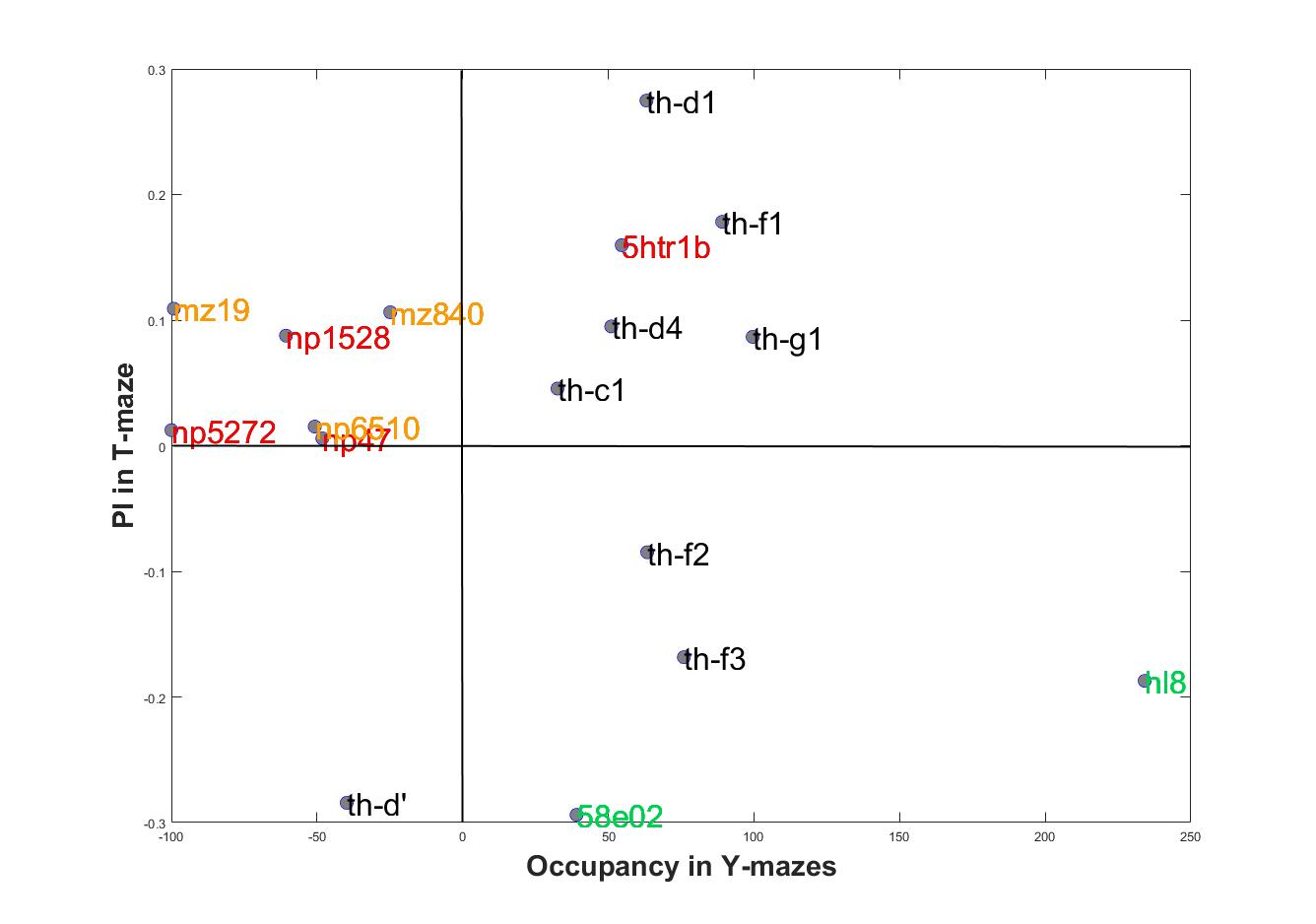
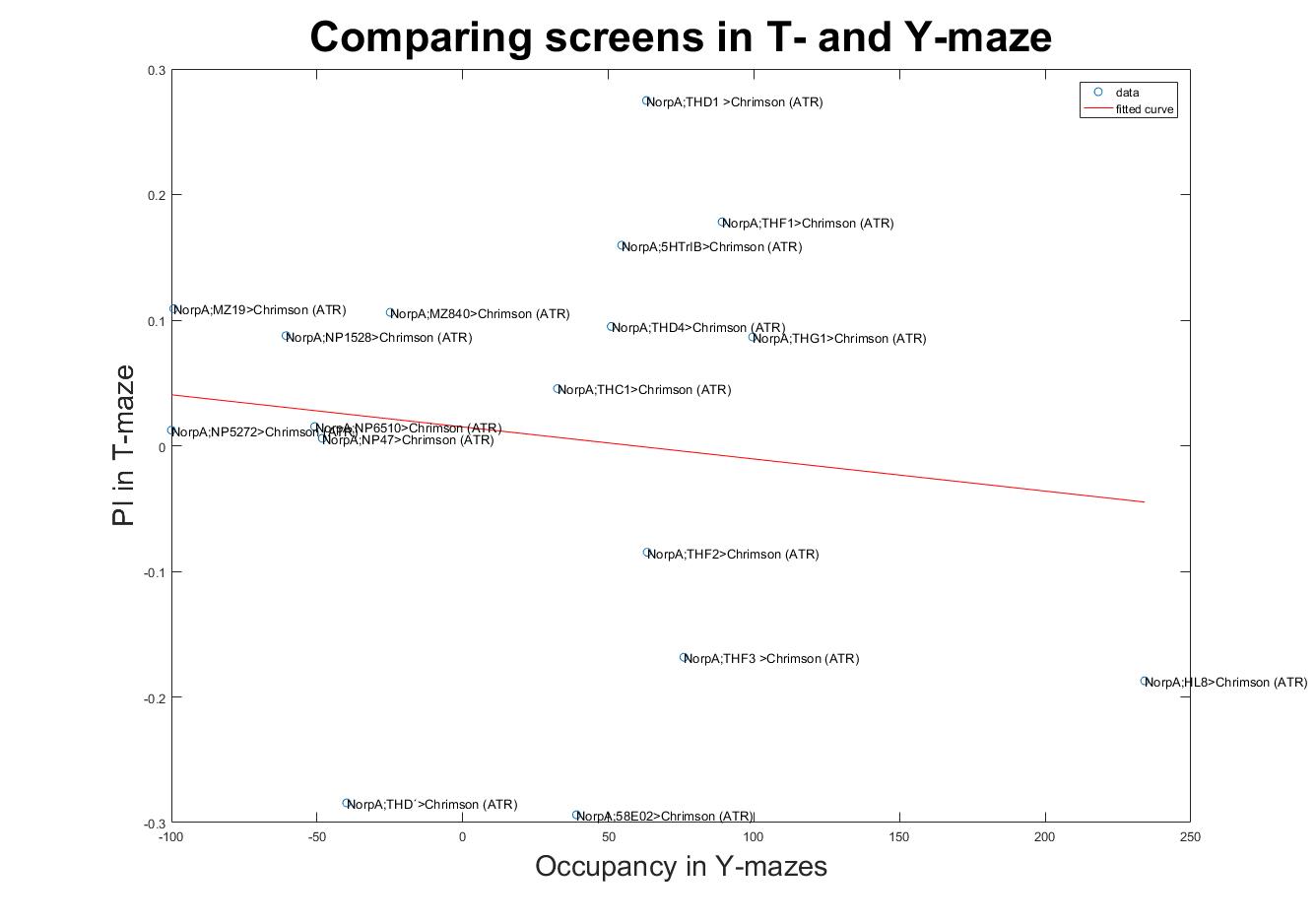 Comparing scores in both setups show some lines that match their scores and other lines that have opposite scores. This filters the ones that are context dependent to the ones that show a context independent reinforcement valence.
Comparing scores in both setups show some lines that match their scores and other lines that have opposite scores. This filters the ones that are context dependent to the ones that show a context independent reinforcement valence.
Category: neuronal activation, Operant reinforcment, Optogenetics | No Comments
Optimizing the Joystick with Gr28bd+TrpA1>Chrimson
on Monday, May 29th, 2017 12:58 | by Christian Rohrsen
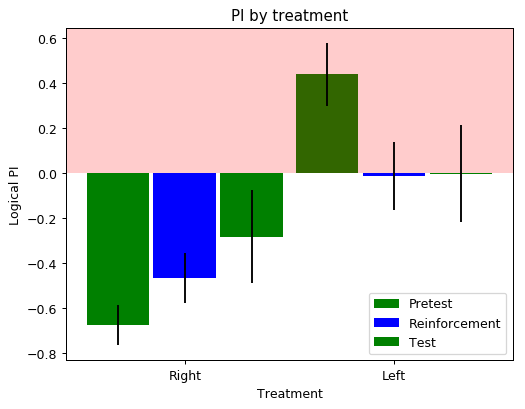 It seems that there is something more there. There were 5 batches of 3 flies per side reinforced. I put the intensity higher than ever before. I will try to get the maximum intensity for the next experiment and see what it looks like.
It seems that there is something more there. There were 5 batches of 3 flies per side reinforced. I put the intensity higher than ever before. I will try to get the maximum intensity for the next experiment and see what it looks like.
Category: neuronal activation, Operant reinforcment, Optogenetics, python code | No Comments
Looking for the phenotype in Gr28bd+TrpA1>Chrimson
on Monday, May 22nd, 2017 3:01 | by Christian Rohrsen
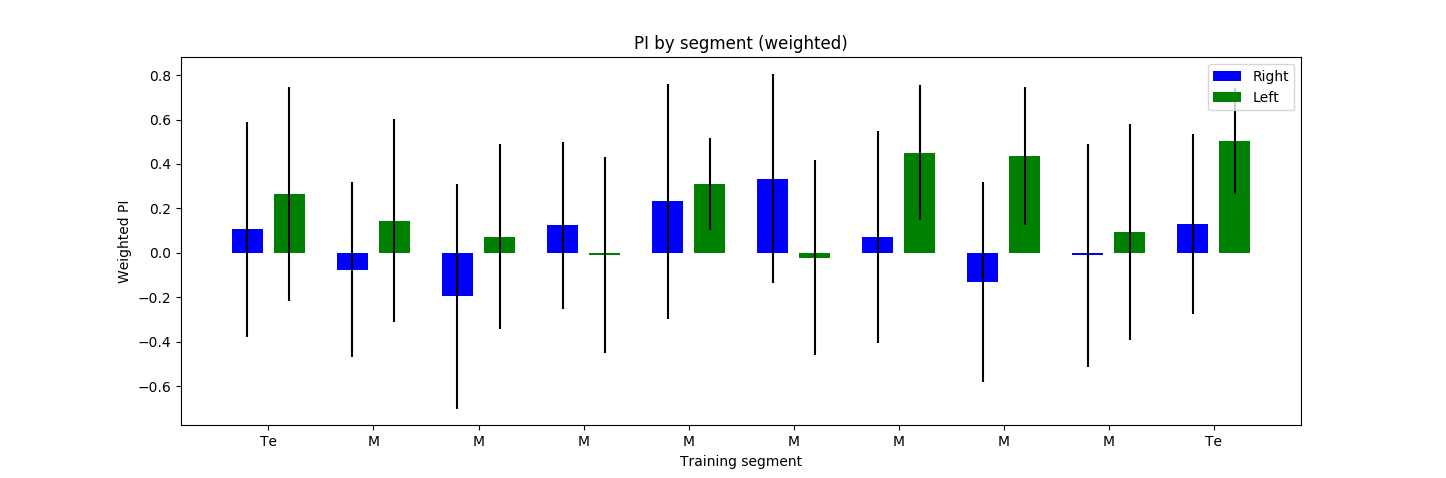
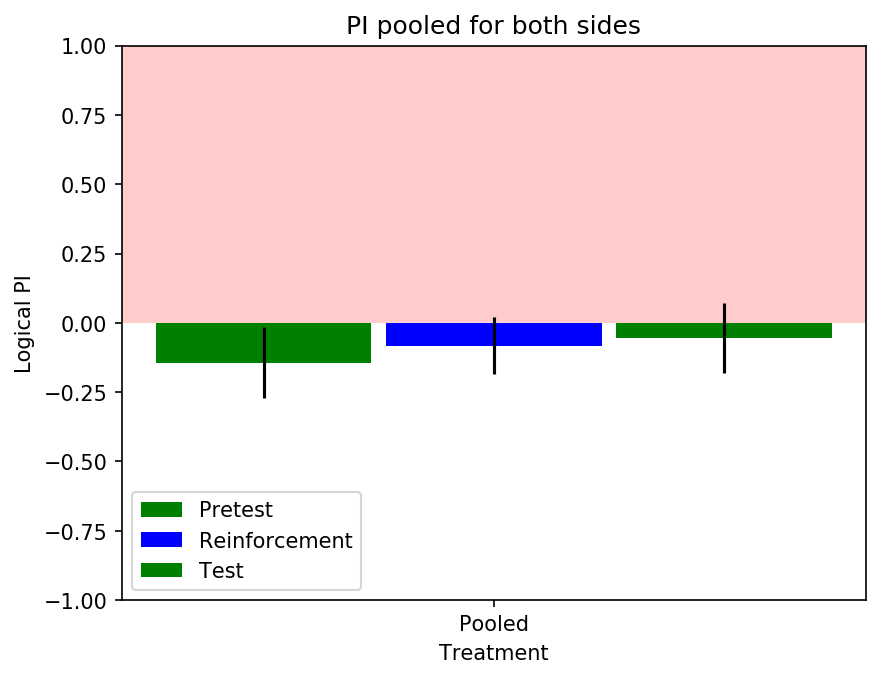
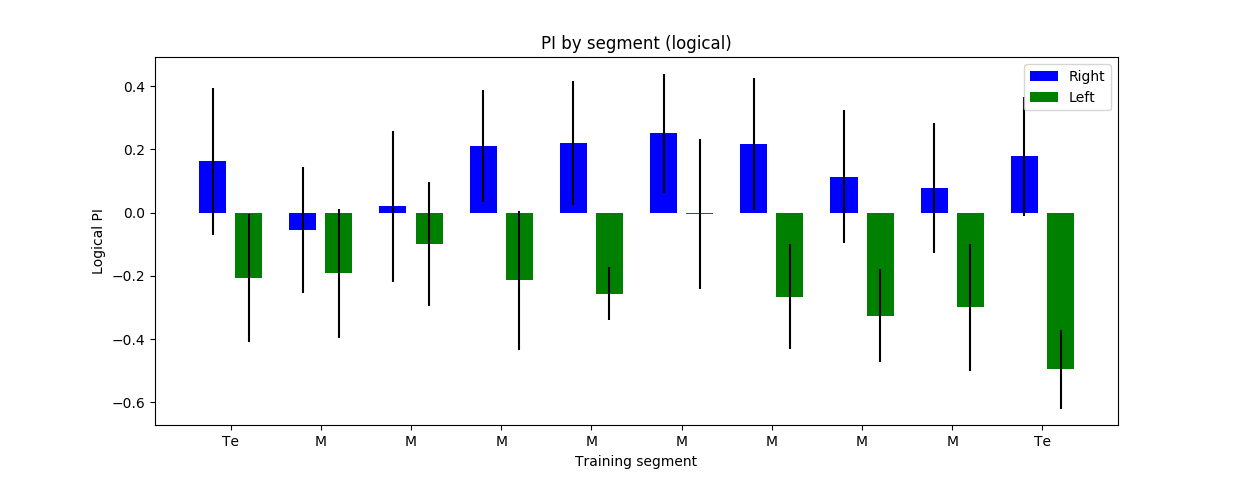
I have decreased the light intensity to around half of what it previously was. The result do not show any clear phenotype. The ‘logical PI’ looks kind of inverted because I actually have plotted it so that positive means light, negative means no light. A total of 30 flies were tested: some of them with right, other with left reinforcement
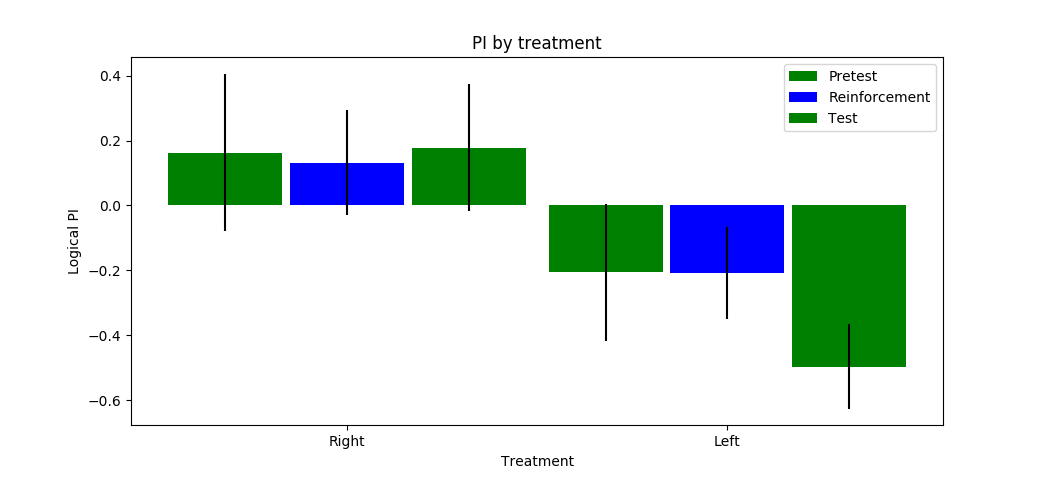
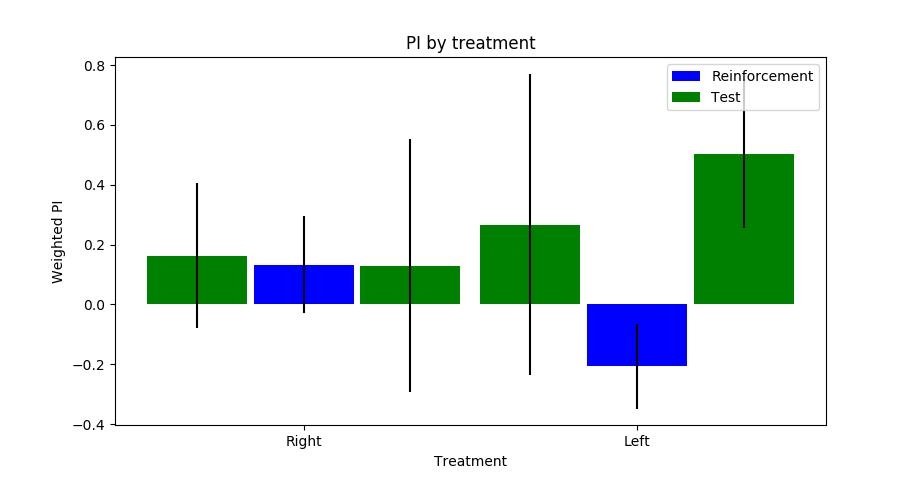
Category: neuronal activation, Operant reinforcment, Optogenetics, python code | No Comments