I) Molecular part
on Monday, July 28th, 2025 1:51 | by Julia Schulz
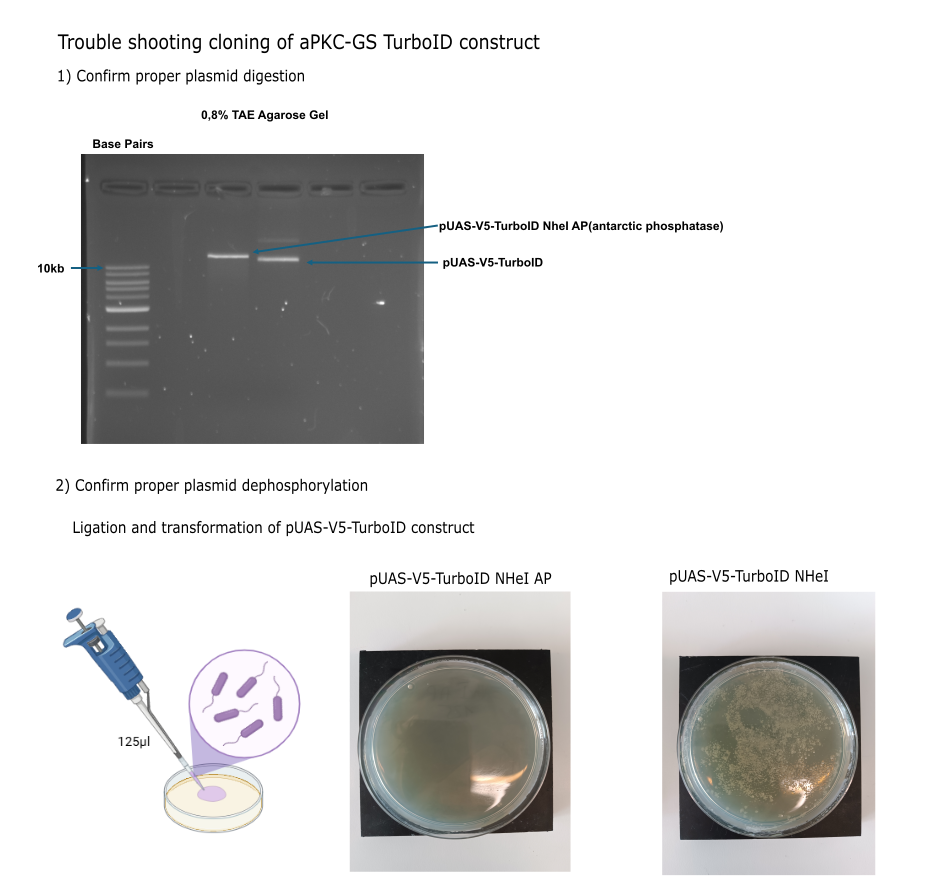
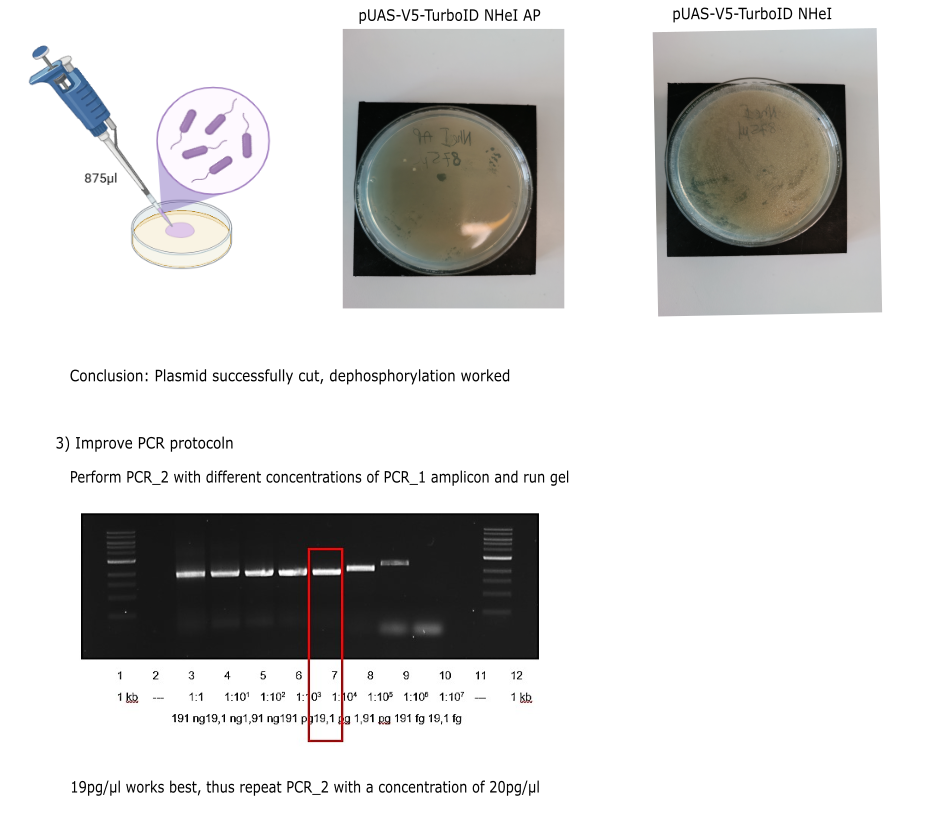

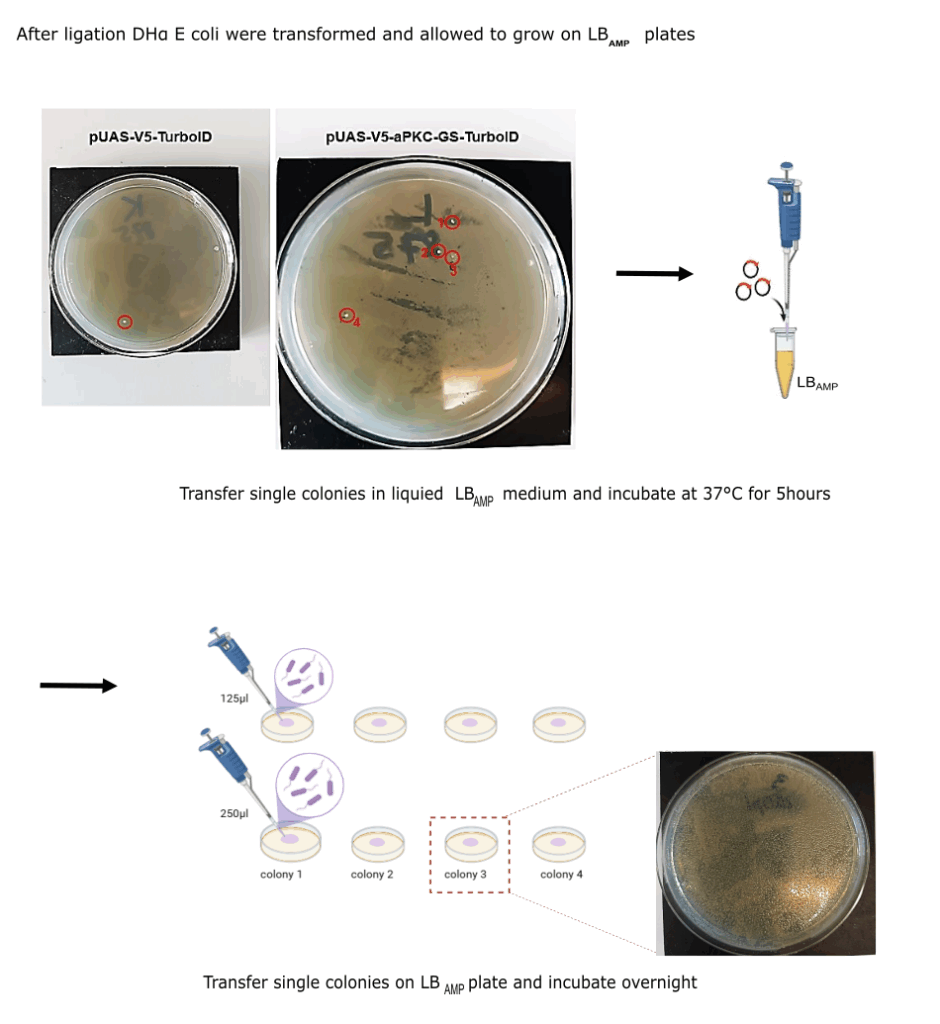
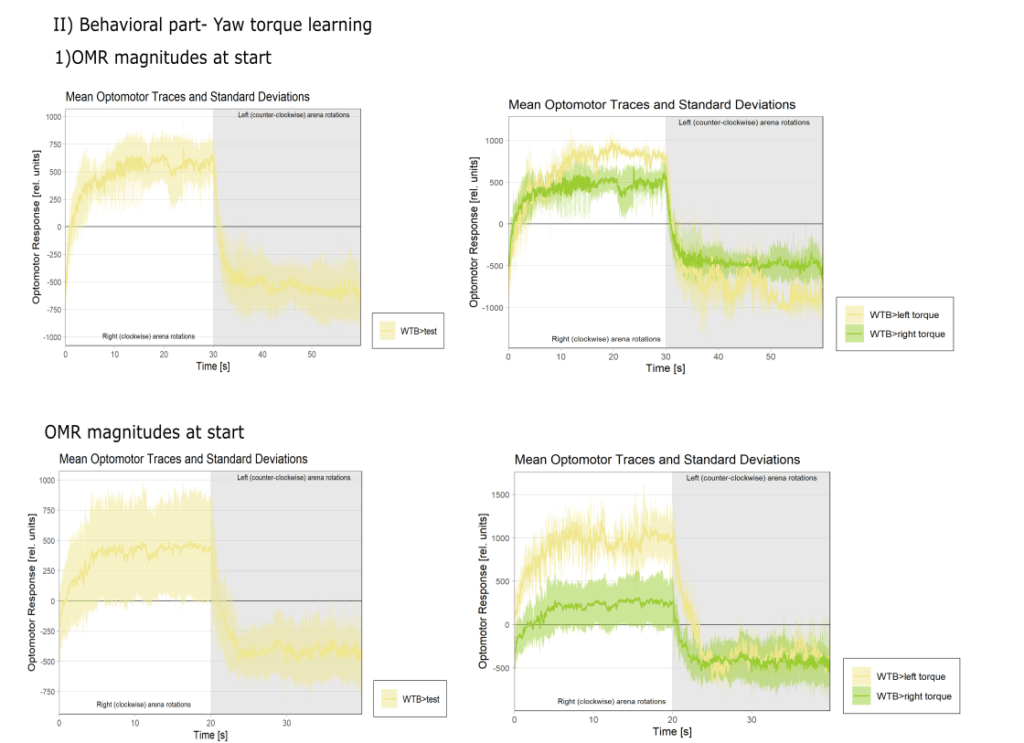


Category: Operant learning, operant self-learning | No Comments
on Monday, July 14th, 2025 1:56 | by Julia Schulz




Category: operant self-learning, Optogenetics | No Comments
Yt learning_positioning effect
on Monday, June 30th, 2025 11:47 | by Julia Schulz
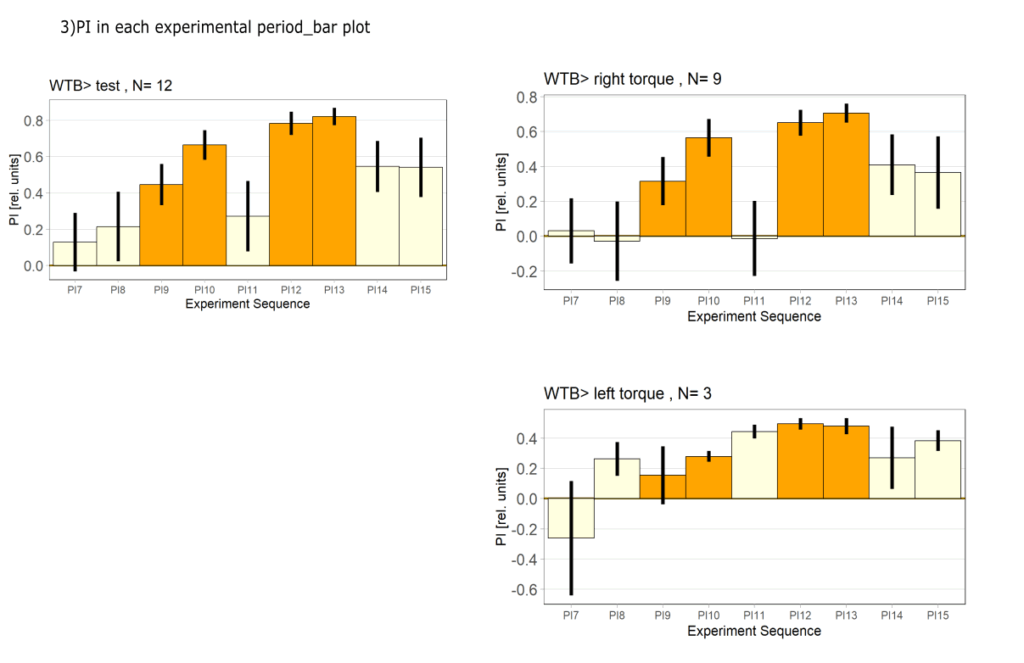
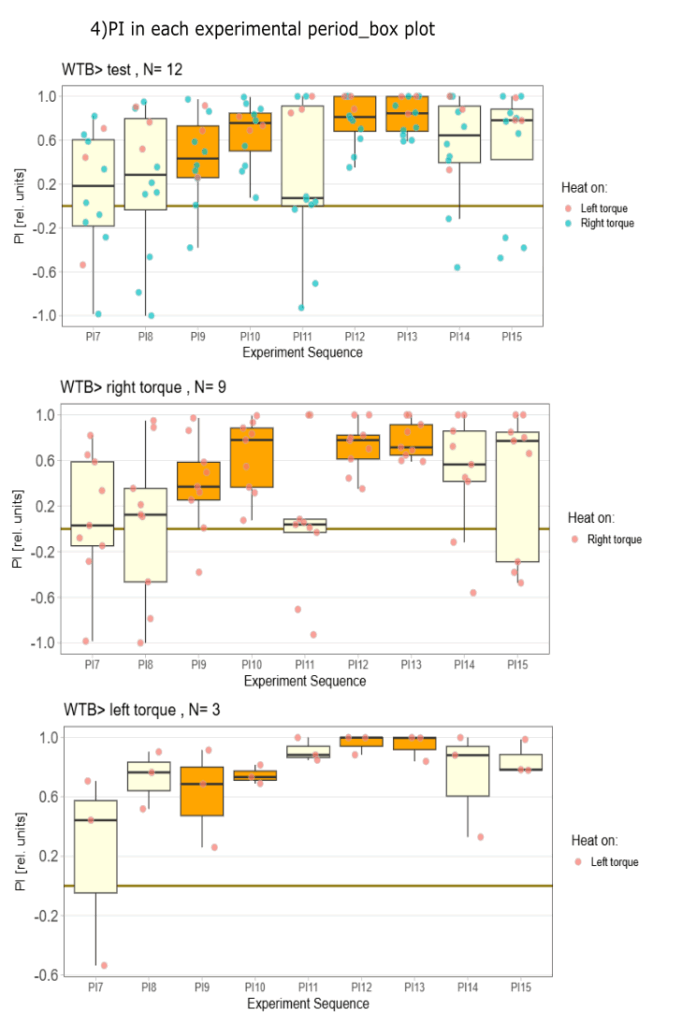
side bias can be compensated for by changing the positioning of the flies within the copper clamp from the medial to the lateral side facing away from the wall.
Category: Operant learning, operant self-learning, Uncategorized | No Comments
YT learning_trouble shooting_body orientation
on Monday, June 23rd, 2025 1:39 | by Julia Schulz
Category: Operant learning, operant self-learning | No Comments
Yt learning_trouble shooting
on Monday, June 23rd, 2025 1:37 | by Julia Schulz
Category: Operant learning, operant self-learning | No Comments
Data sets pooled
on Thursday, May 8th, 2025 8:01 | by Björn Brembs
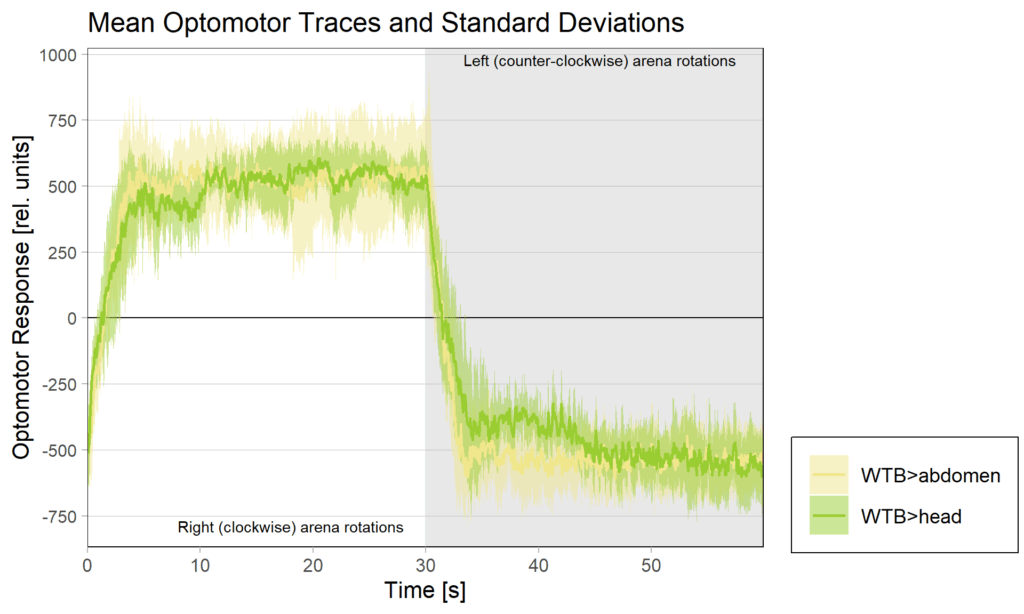
I managed to write some code that shortens Tina’s optomotor periods and then adjusted the metadata between our experiments so that the script would run without any errors. So this is what our combined tests of expressing the foraging RNAi in all neurons looks like:

The full dataset is also available on the UR publication server.
Category: Operant learning, operant self-learning, PKG, Rover/Sitter | No Comments
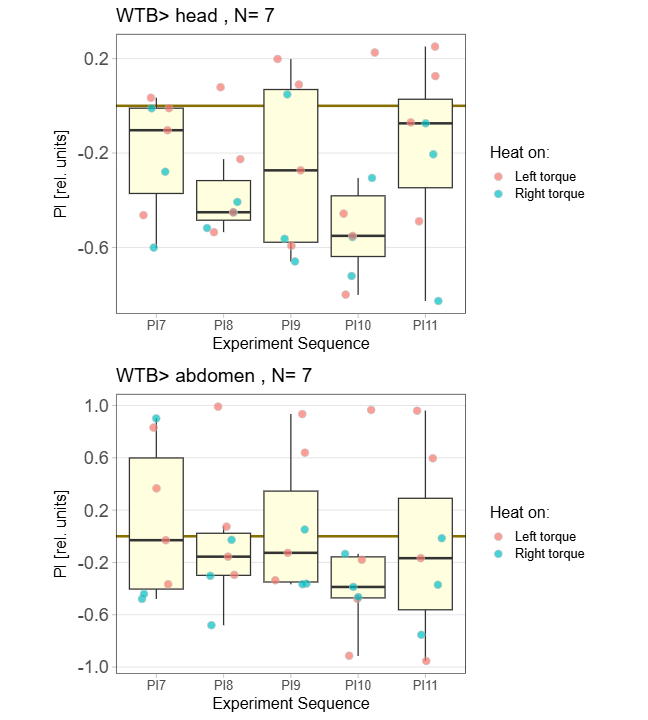
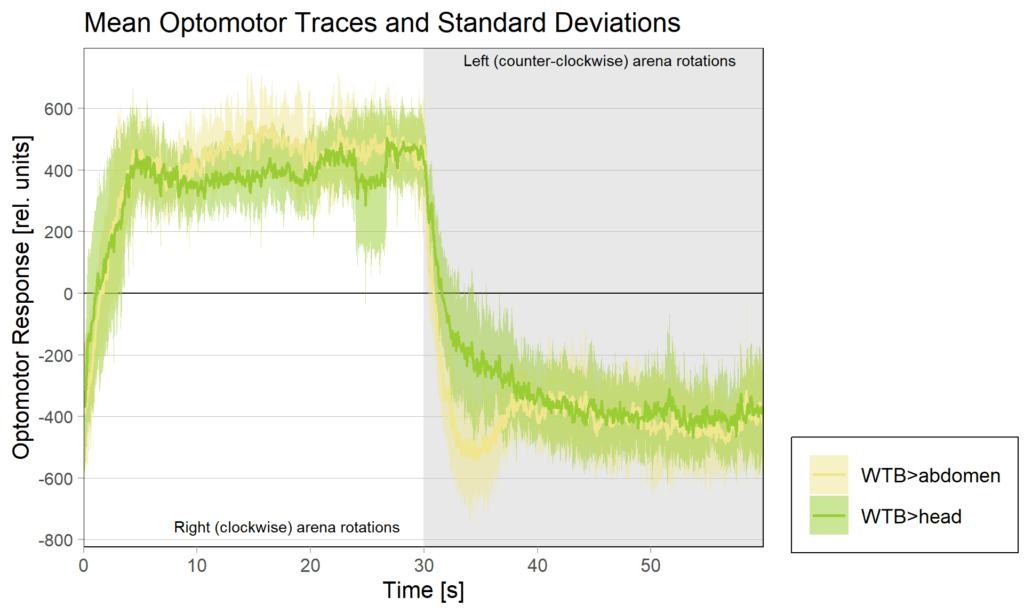
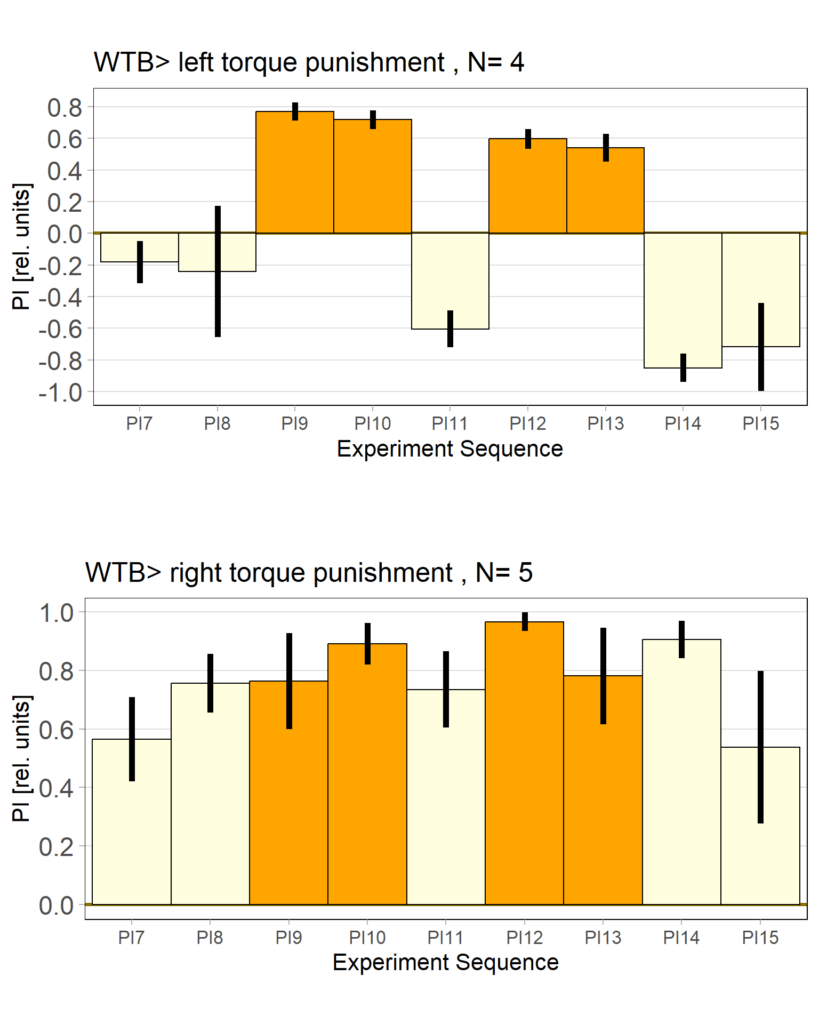
yaw torque measurement_Dtc 20%
on Monday, April 14th, 2025 1:20 | by Julia Schulz
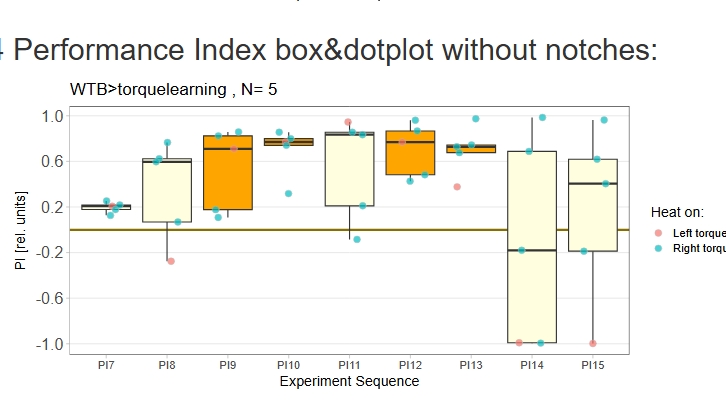
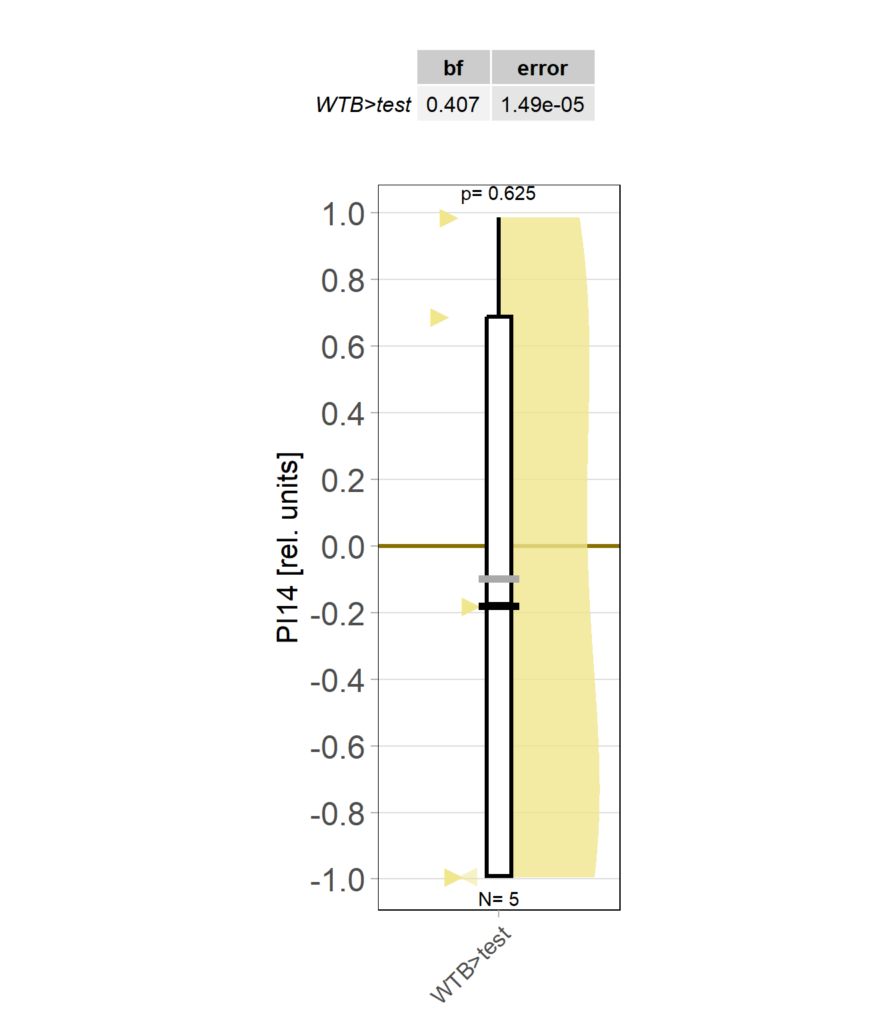
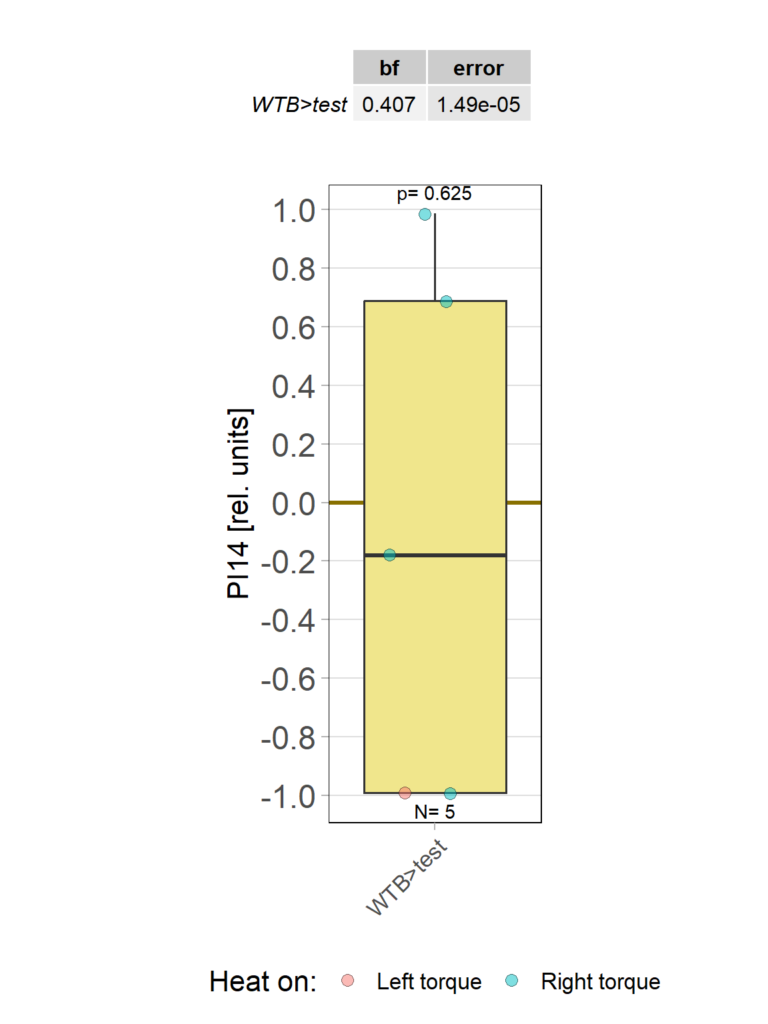
Category: operant self-learning, Optomotor response | No Comments
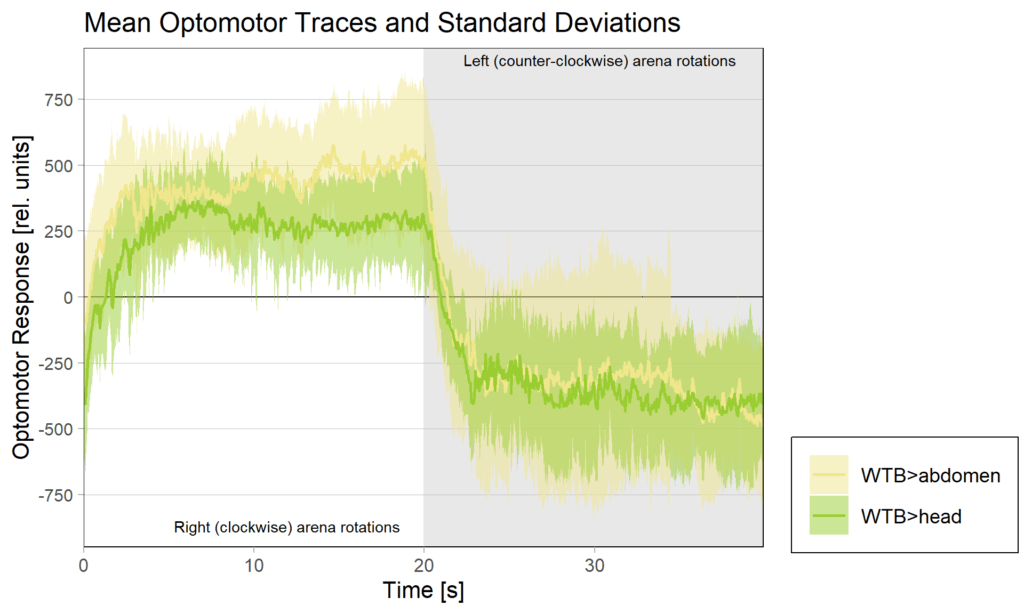
operant self learning_Dtc 40
on Monday, April 14th, 2025 1:13 | by Julia Schulz
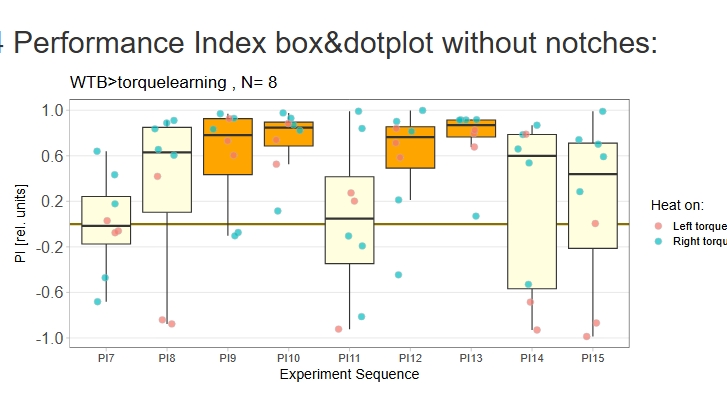
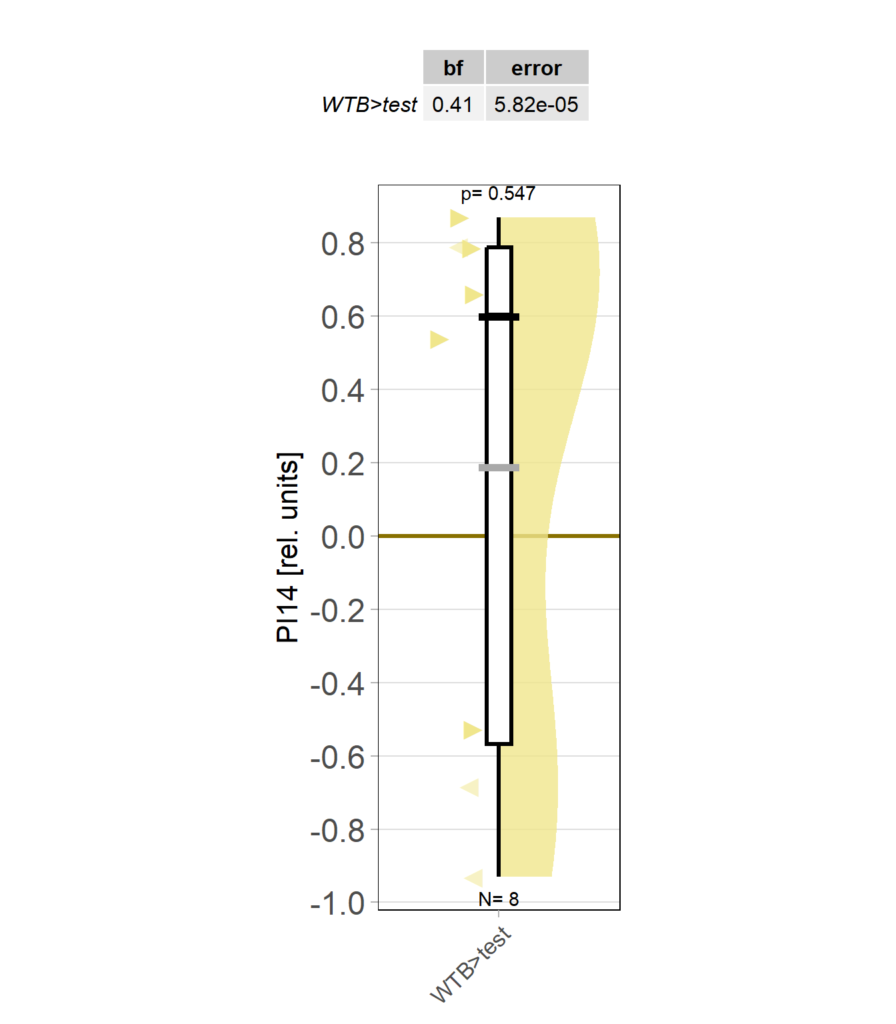
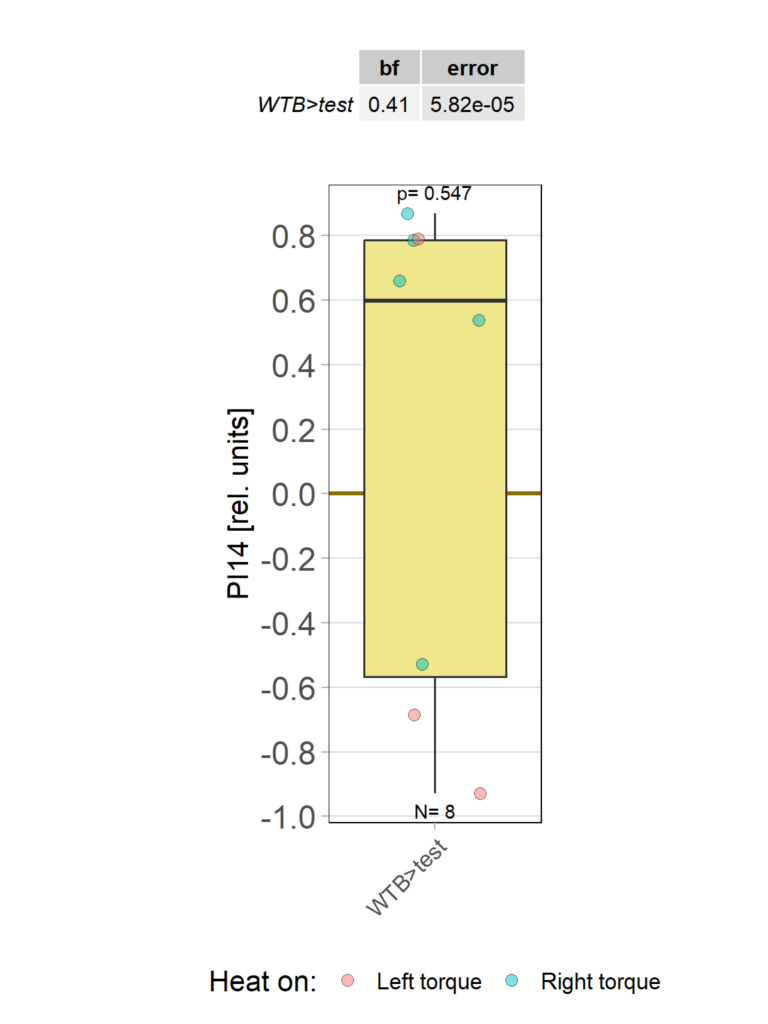
Category: operant self-learning, Optomotor response | No Comments
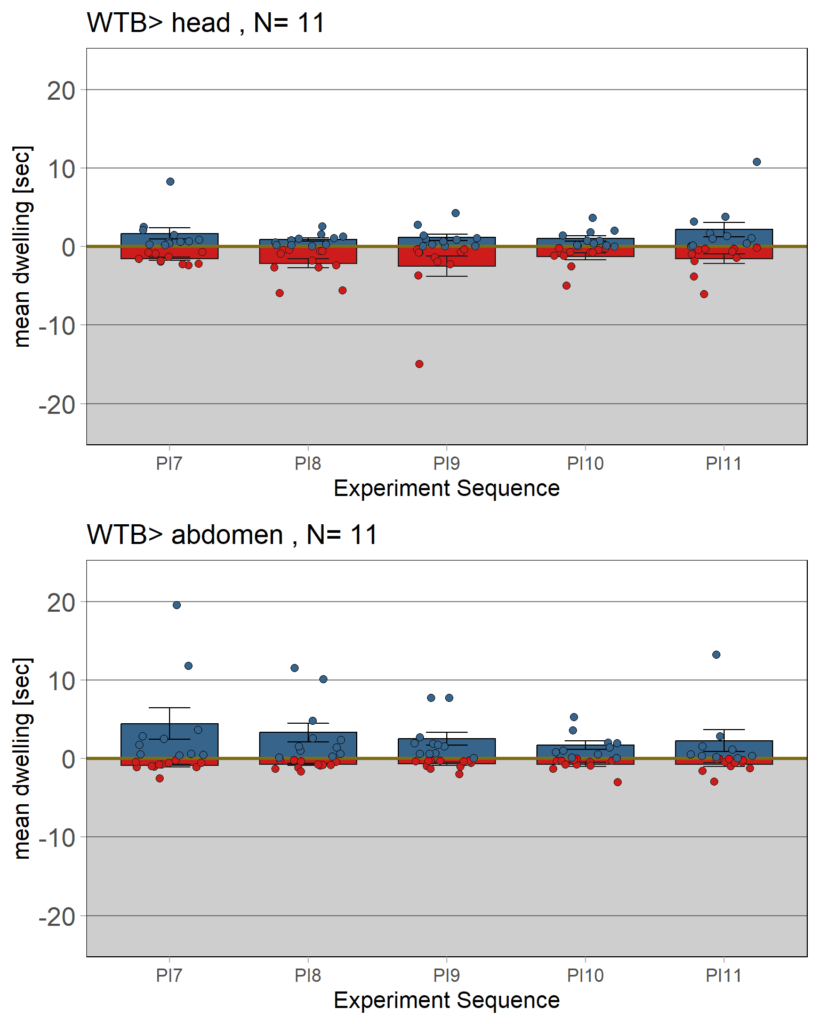
yaw torque learning_Dtc_40%
on Monday, March 31st, 2025 10:30 | by Julia Schulz
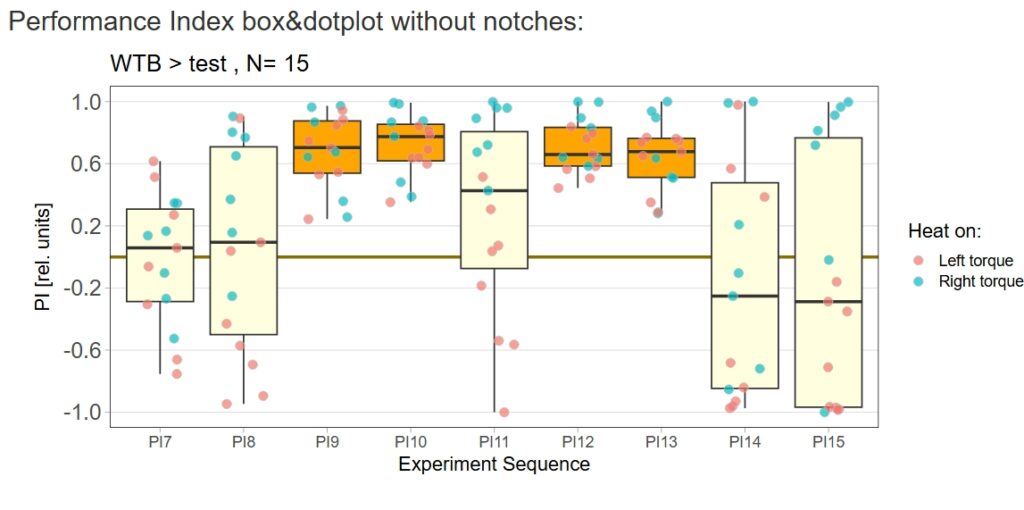
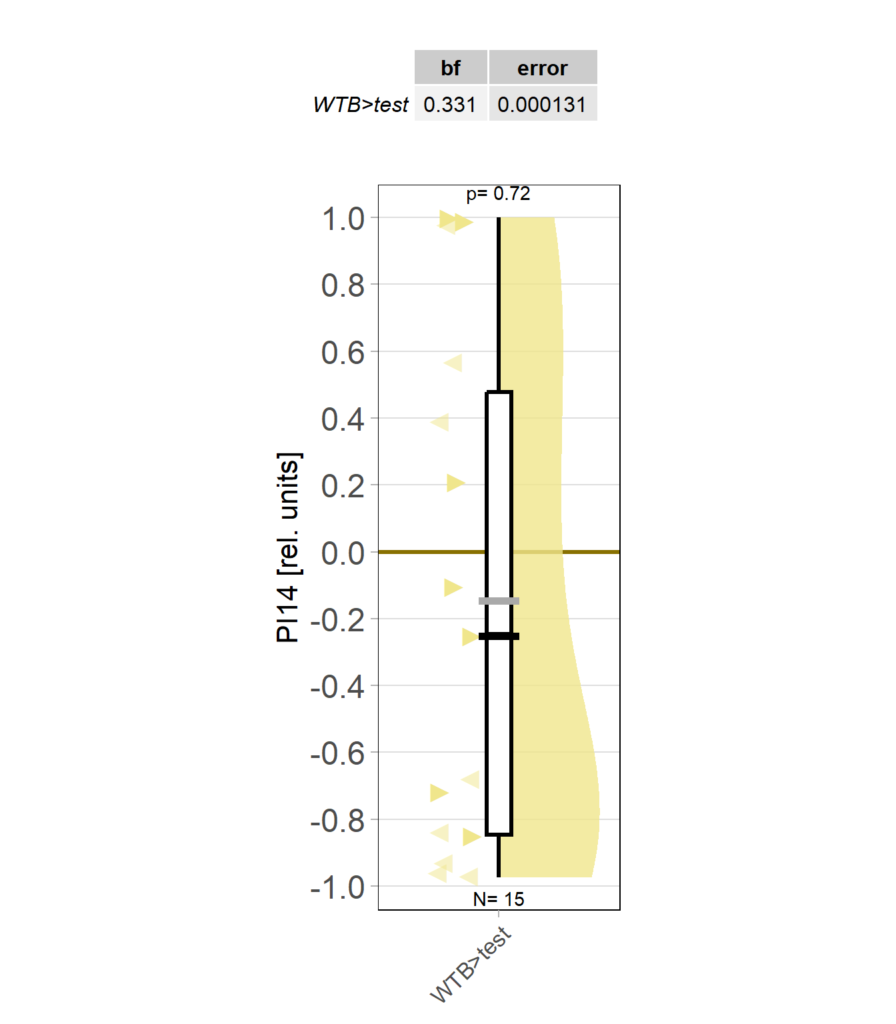
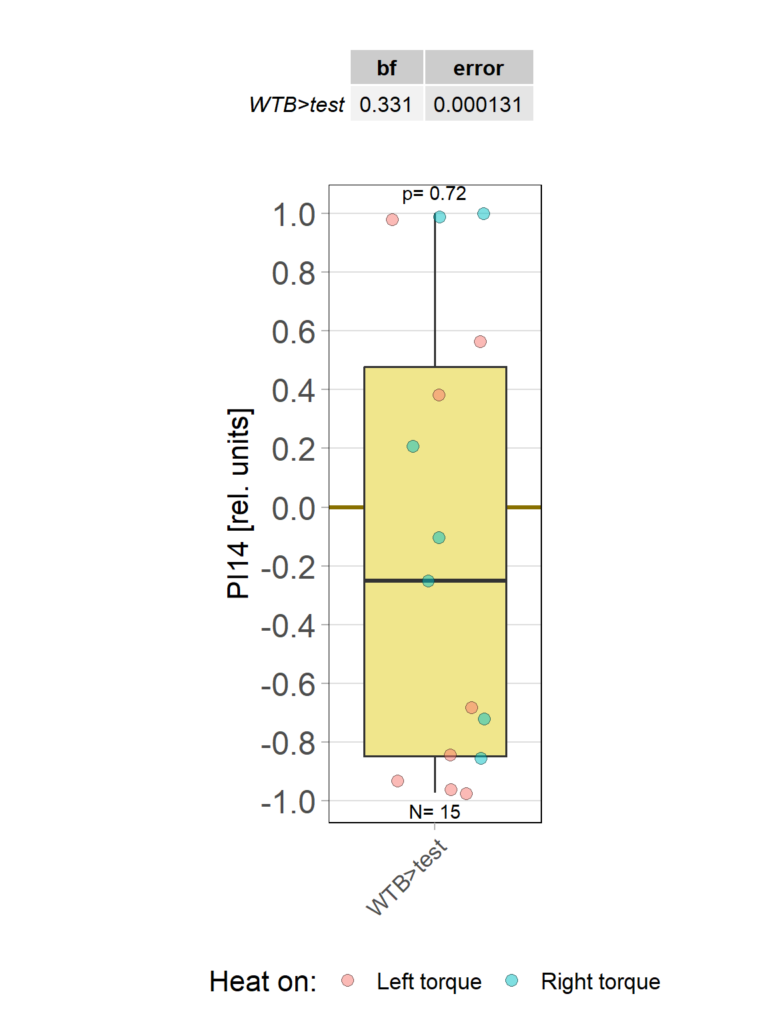
Category: Operant learning, operant self-learning | No Comments
Not an unambiguous result
on Friday, March 28th, 2025 3:53 | by Björn Brembs
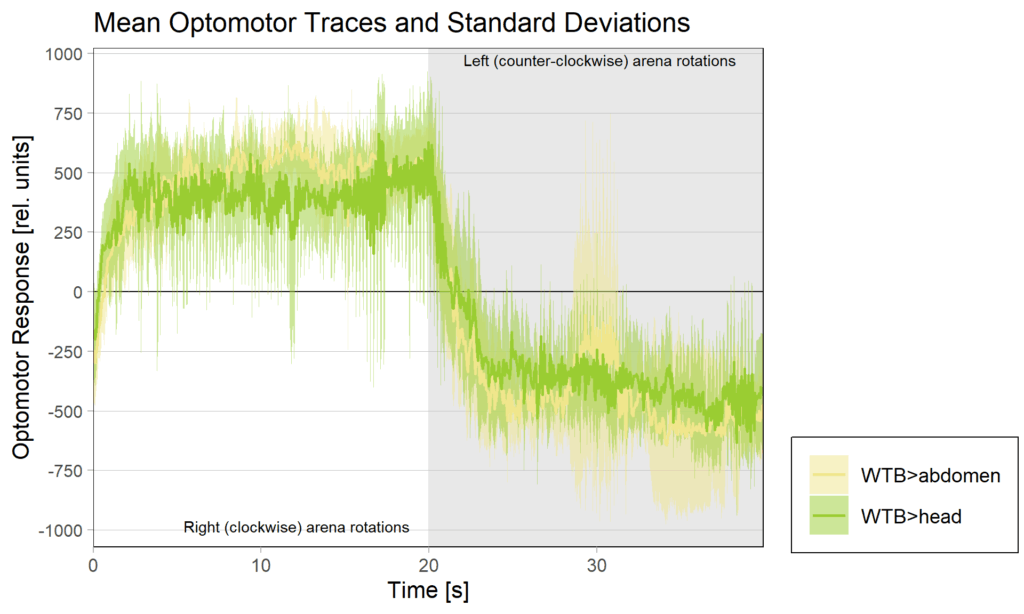
This is the final graph of the experiments knocking down PKG (foraging) in operant self-learning:

The data have been automatically uploaded via the new script and can be found on the University’s publication server.
This was supposed to be a reproduction of Tina’s experiment:

While in my dataset, the knock-down flies are still performing the worst of all three groups, it is not quite as clear-cut as in Tina’s results. Then again, her knock-down flies maybe were slightly too low. Now pooled with mine that were slightly to high, maybe this is what we should expect: a clear zero. We should pool the data and see how it looks.
Category: operant self-learning, PKG | No Comments