NorAUASChrimsonxGr28bdTrpA1_Yellow
on Monday, July 18th, 2022 9:13 | by Melis Öztibet
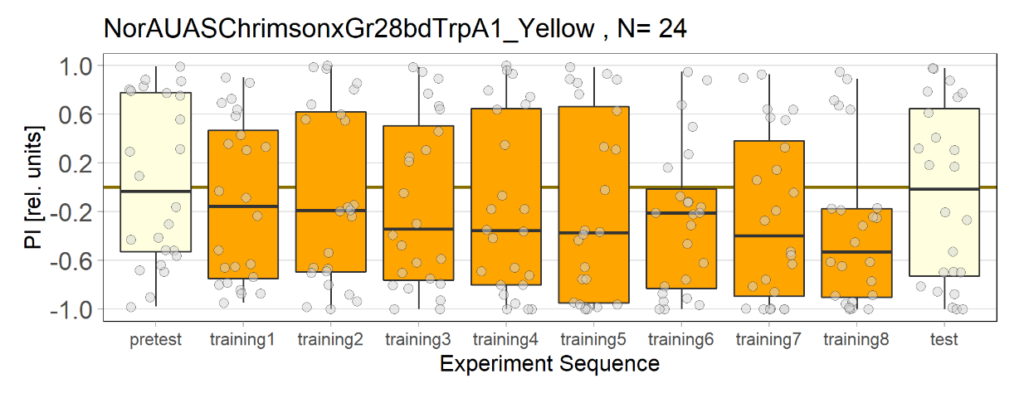

Category: Uncategorized | No Comments
aPKCdelta blind half time experiment
on Monday, May 23rd, 2022 1:19 | by Amelie Hauser
Progress report on the aPKCdelta experiment with half the time (1min period)
Blind experiment with Group A&B; One with Ru486 and one without
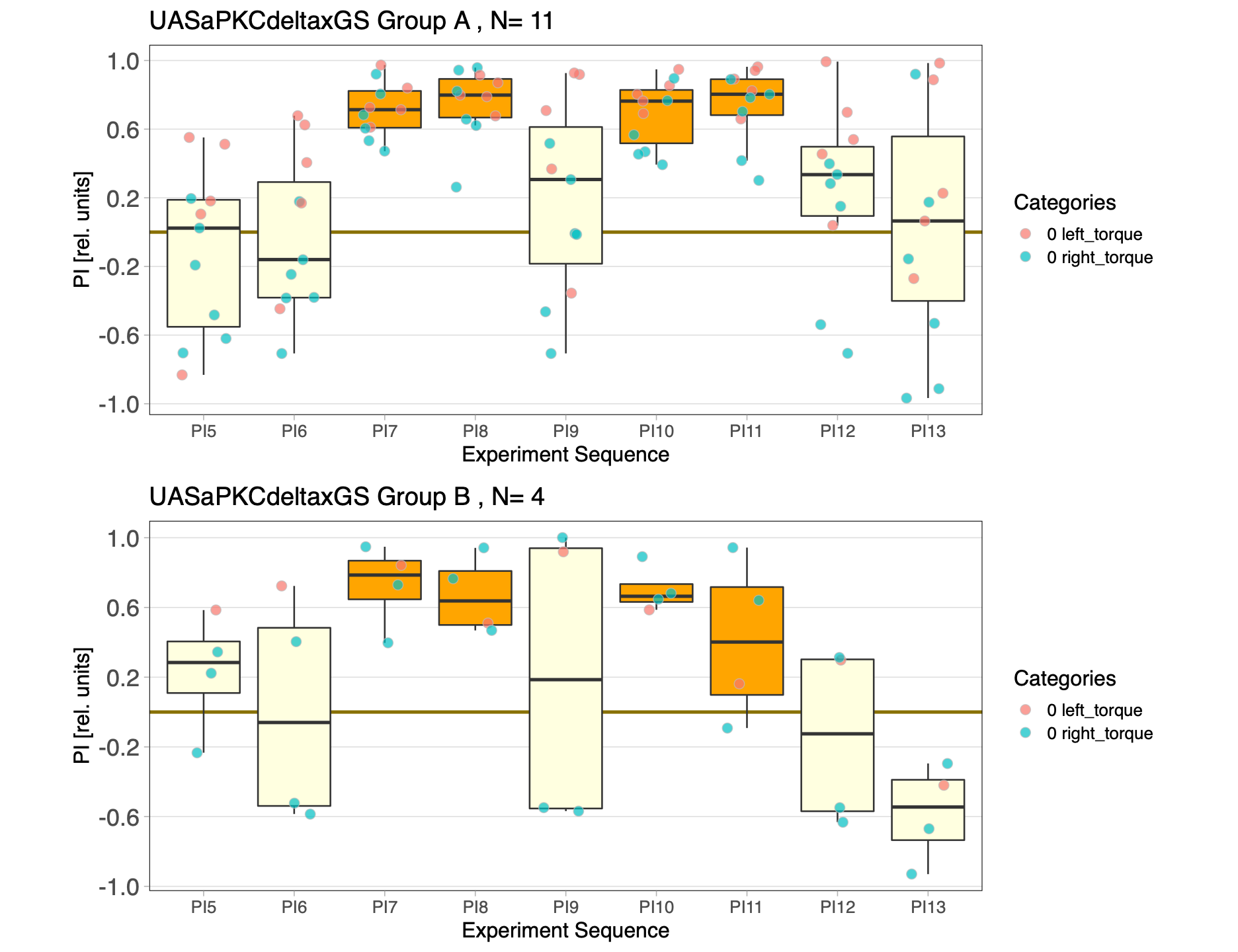
Category: operant self-learning, PKC, Uncategorized | No Comments
Test group for 1 min experiments
on Monday, May 23rd, 2022 1:15 | by Amelie Hauser
Progress report for the test group in the 1 min. period experiments
UASaPKC x GSnsyb control flies
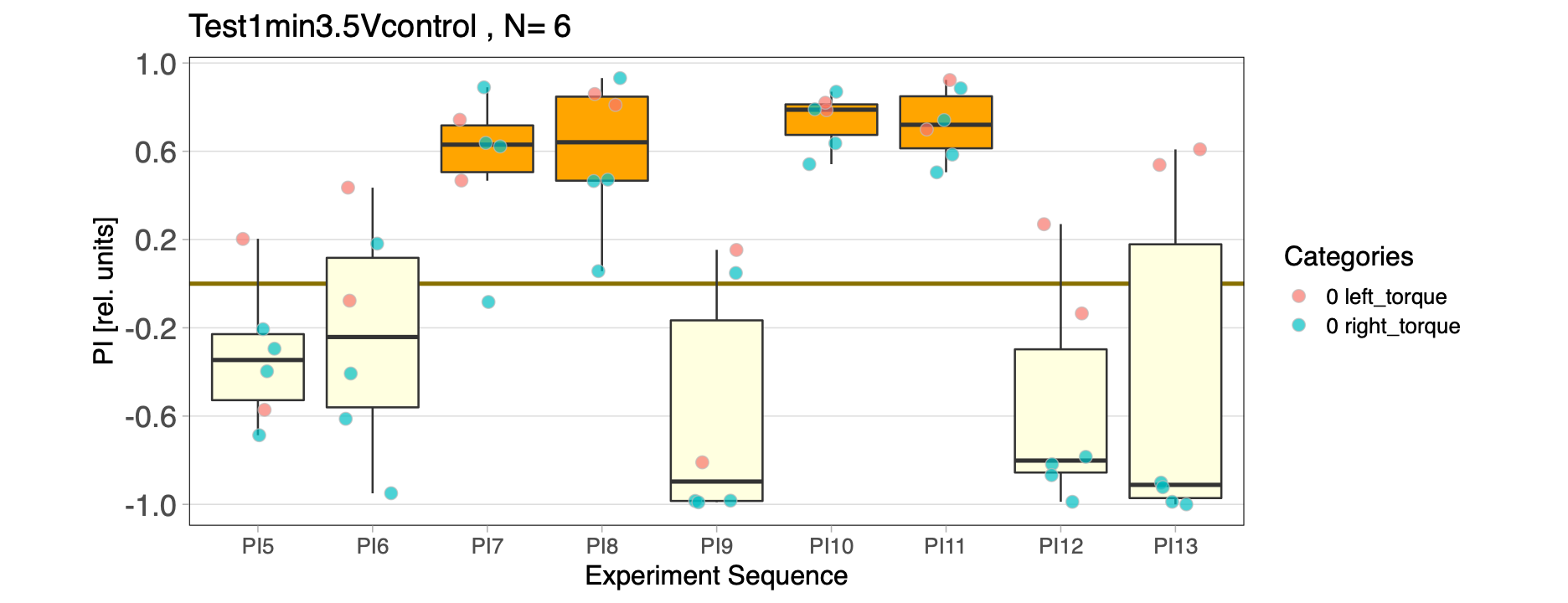
Category: operant self-learning, PKC, Uncategorized | No Comments
Control Groups With and Without ATR
on Monday, May 23rd, 2022 1:13 | by Melis Öztibet
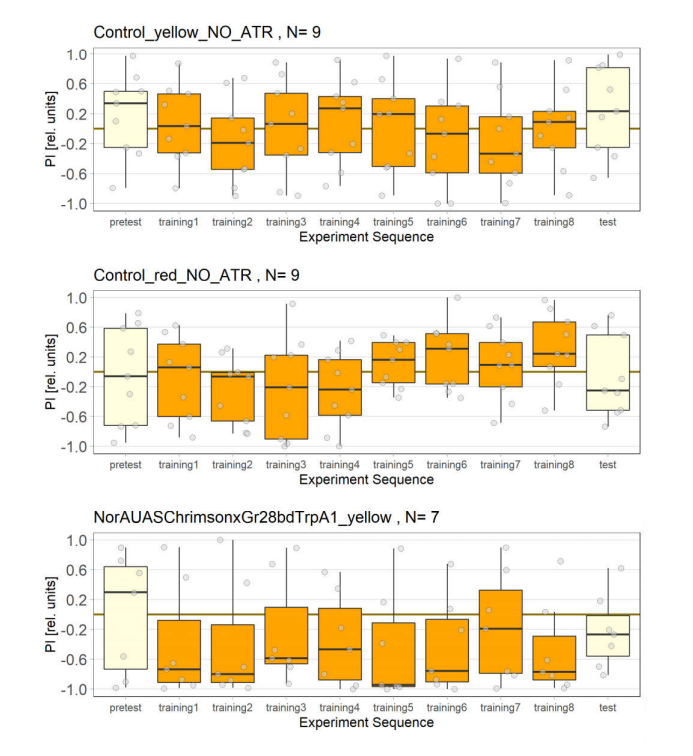
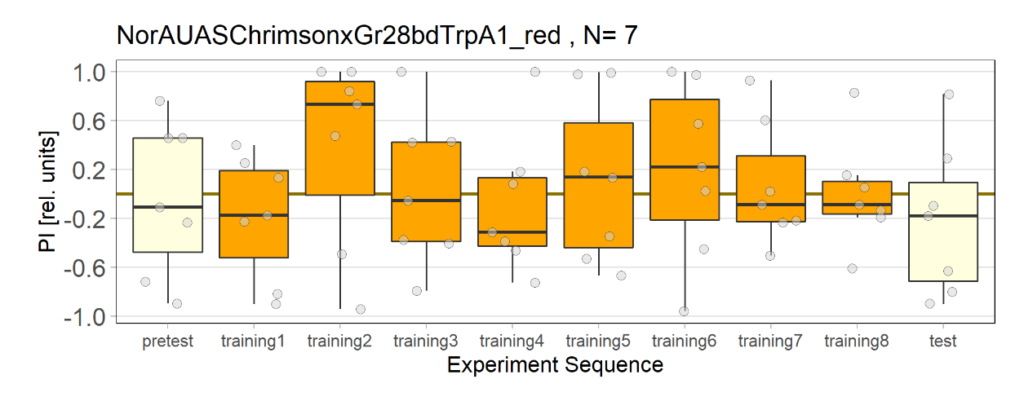
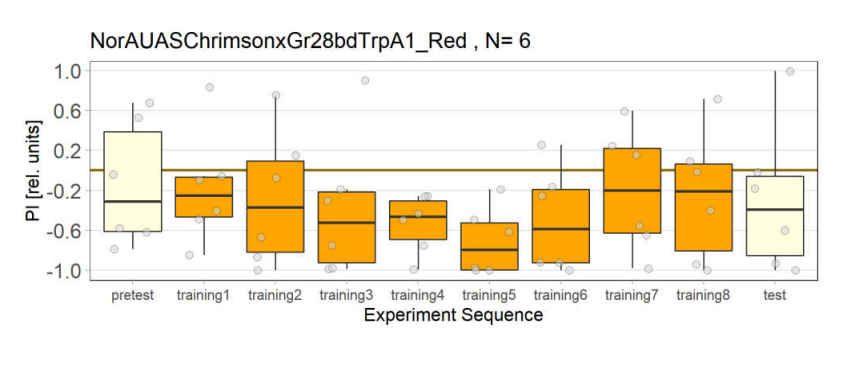
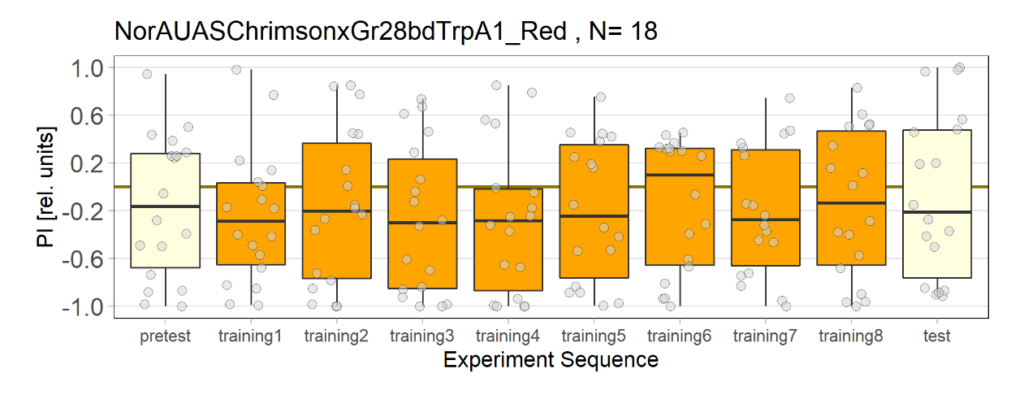
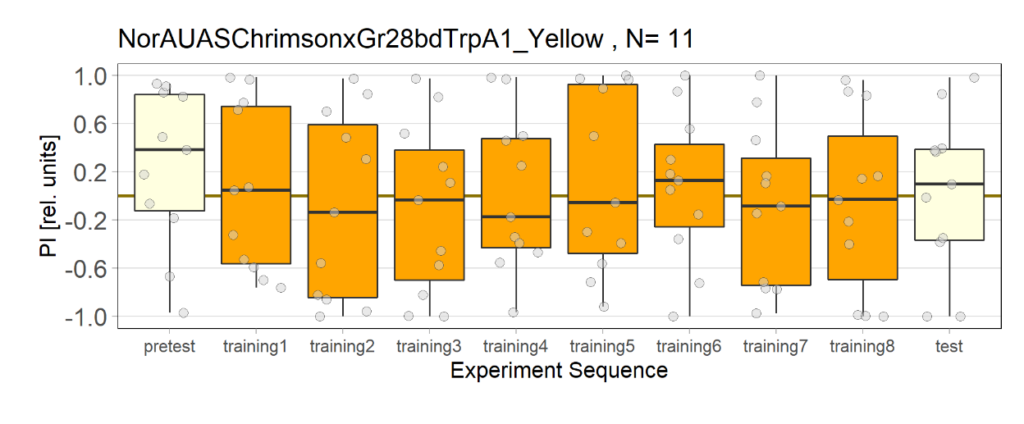
NorAUASChrimsonxGr28bdTrpA1 cross has been used as a control group for joystick experiments. There are 5 different groups for controls.
1st Group: Yellow light, NO ATR
2nd Group: Red light, NO ATR
3rd Group: Yellow light, with ATR
4th Group: Red light, with ATR(2 out of 3 lights have issues)
5th Group: Red light, with ATR(all lambs are working fine for the first time)
6th Group: Red light, with ATR(higher light intensity)
7th Group: Yellow light, with ATR
Category: Uncategorized | No Comments
Baz knockout
on Wednesday, April 20th, 2022 8:20 | by Andreas Ehweiner
Baz was knocked out in all neurons in the adult fly using the gene switch system.
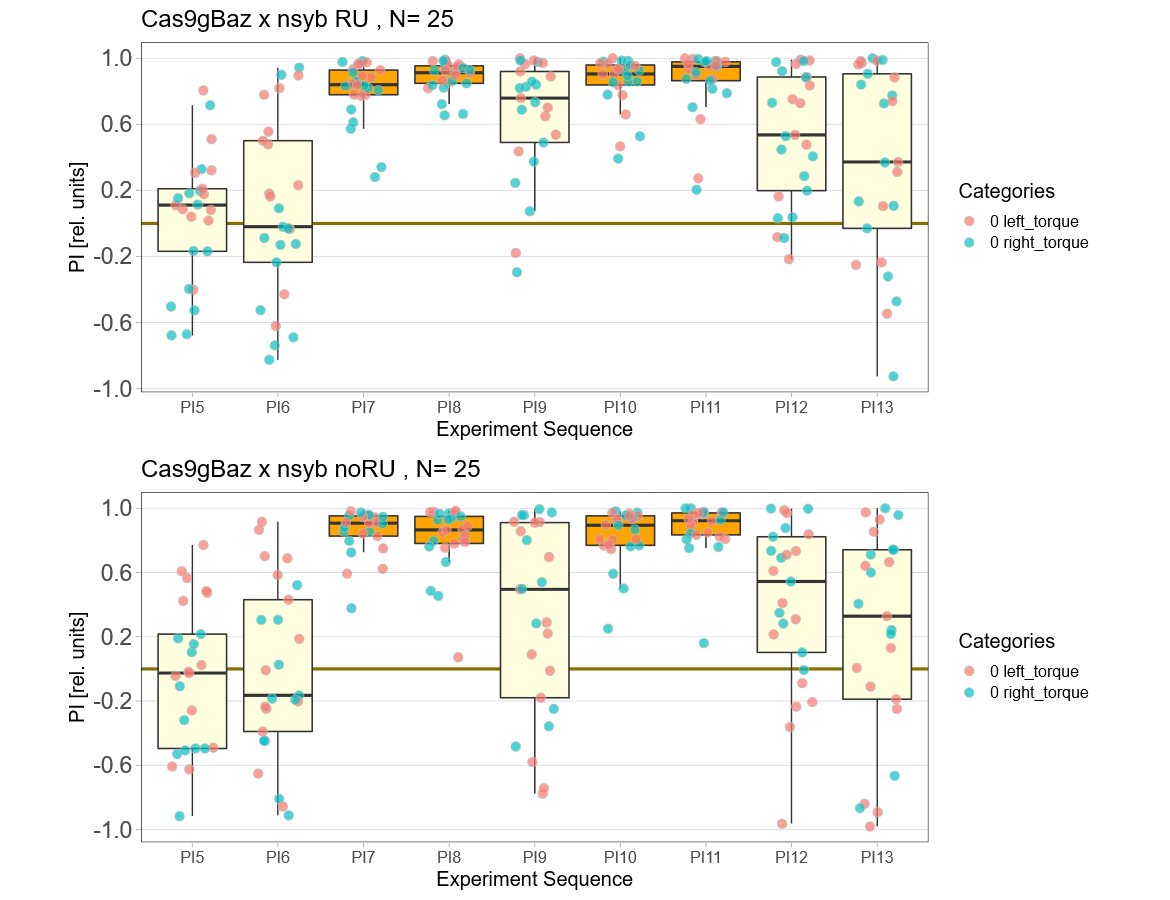
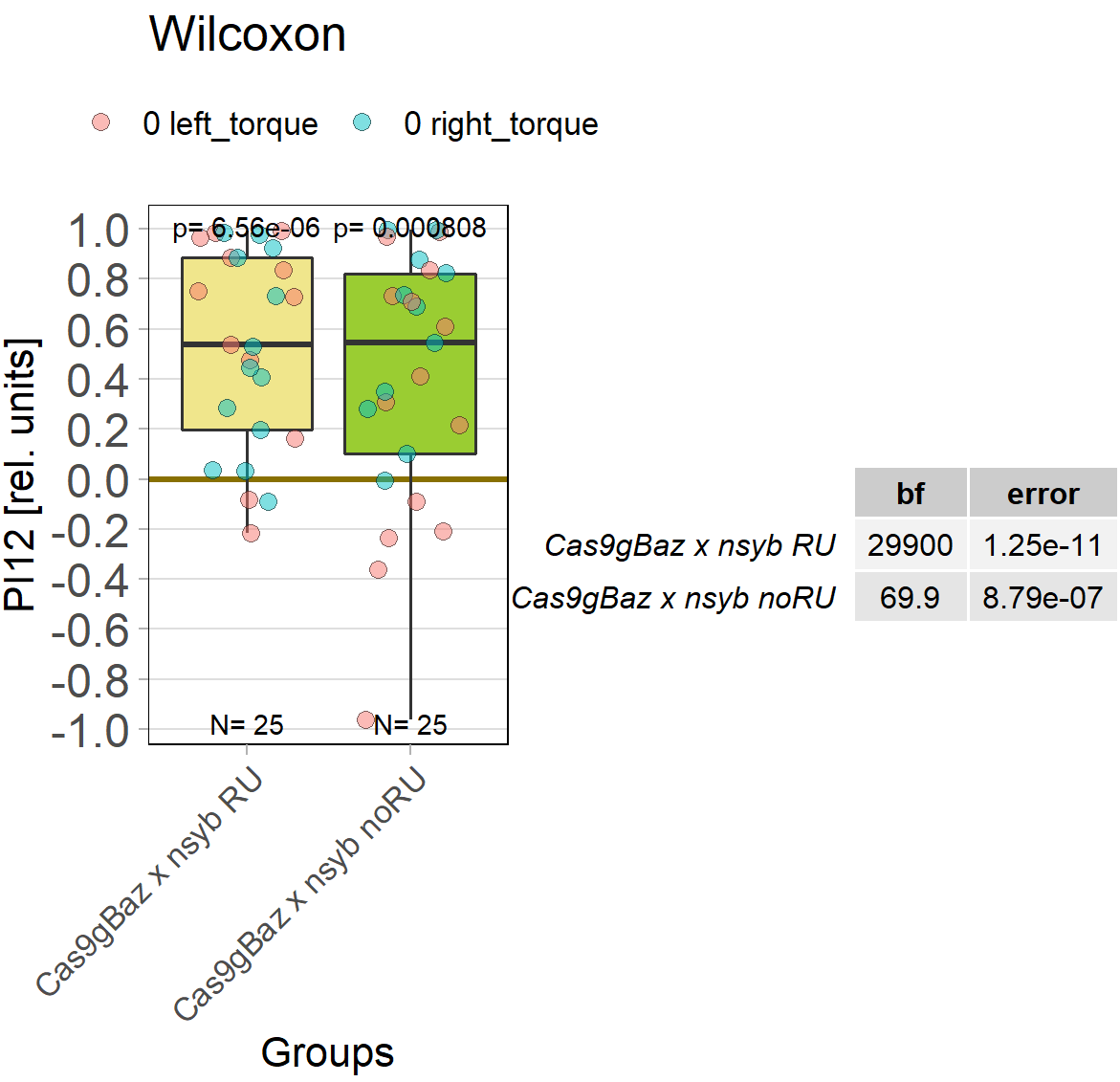
Category: flight, Memory, Operant learning, operant self-learning, Uncategorized | No Comments
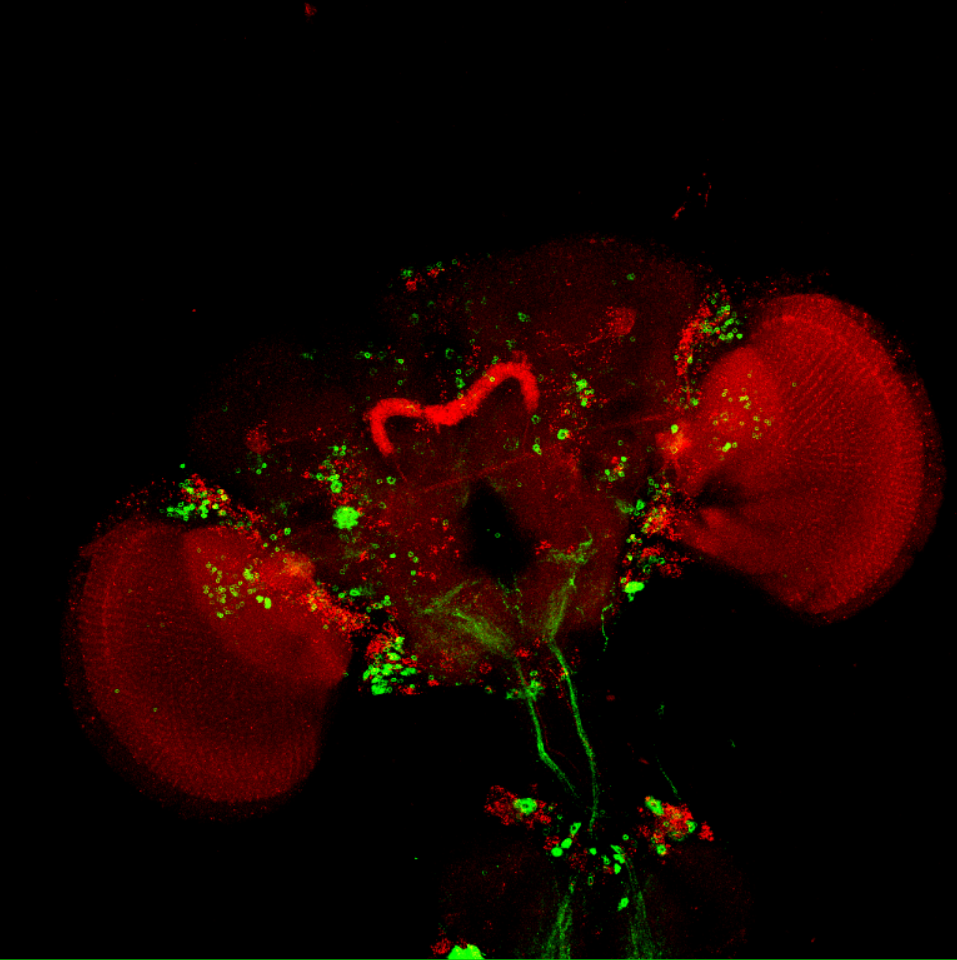
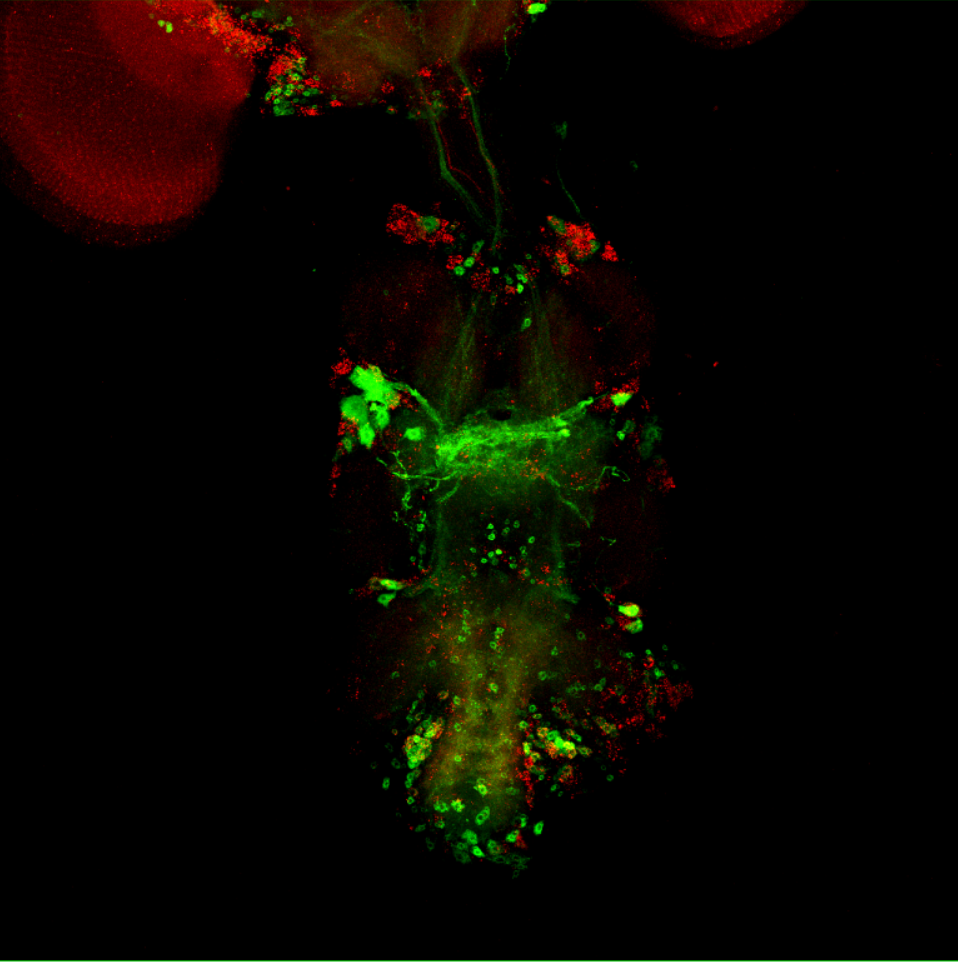
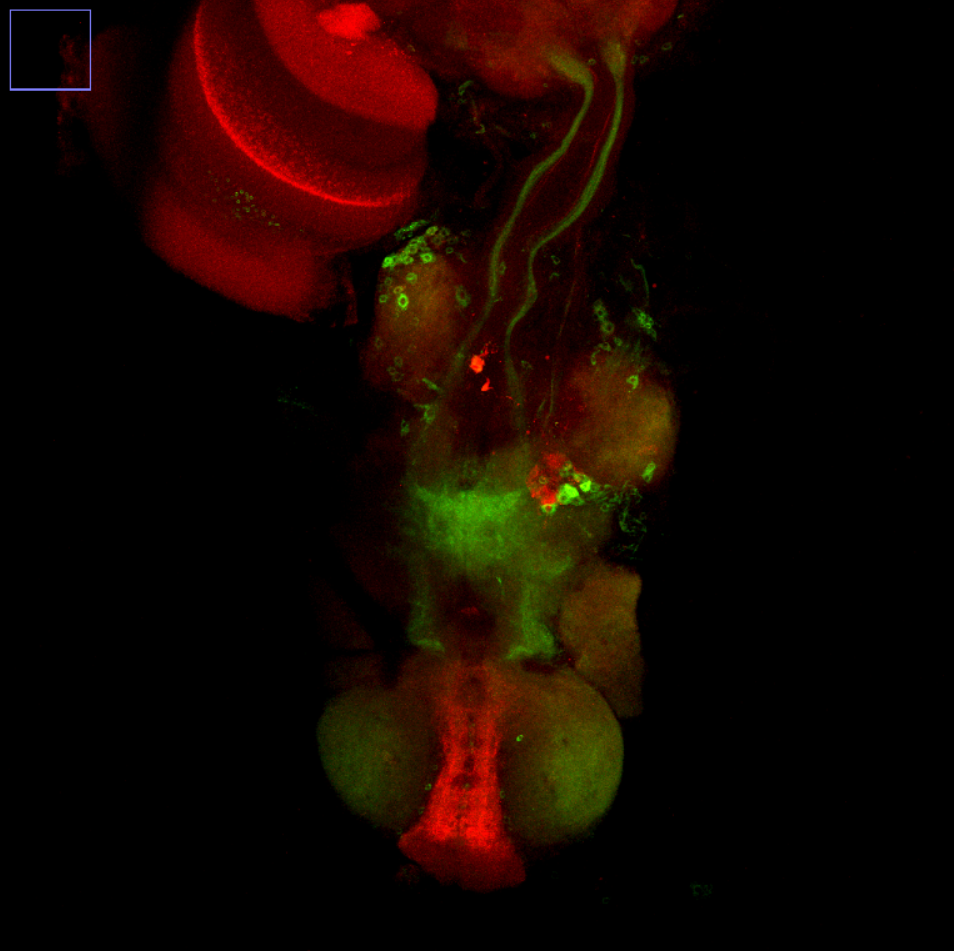
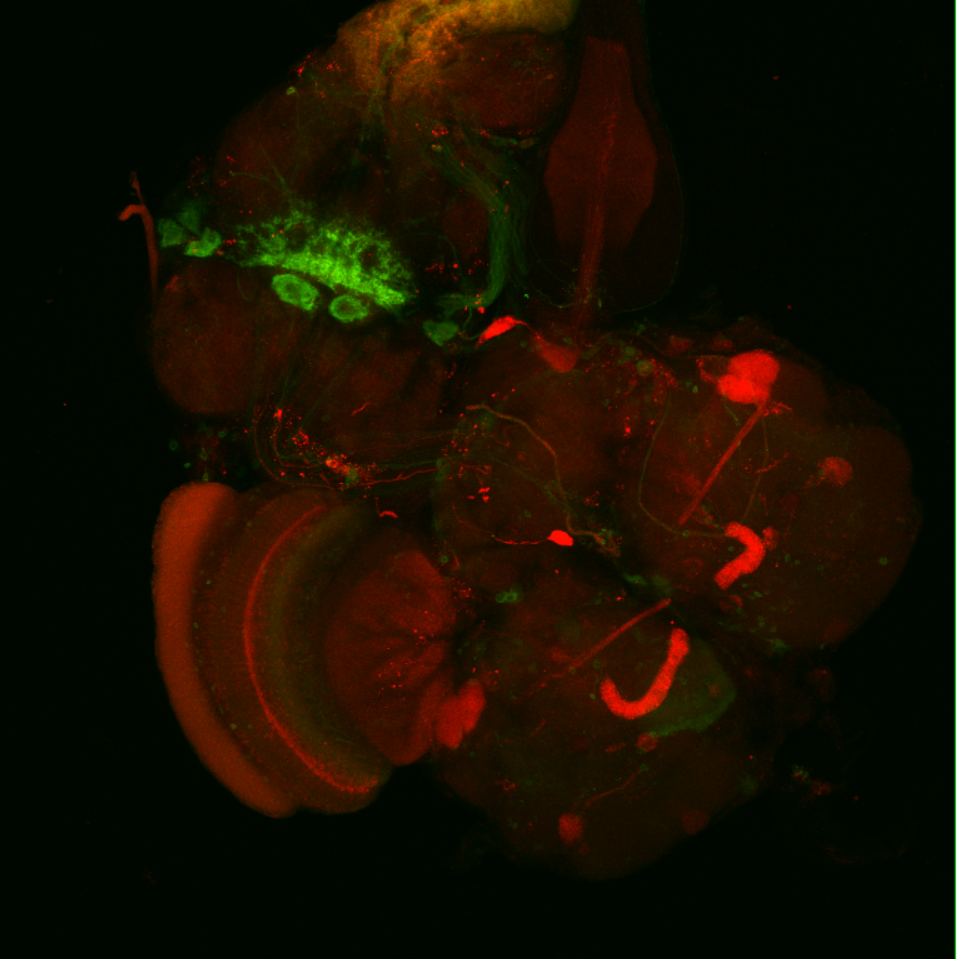
Overlap FoxP – motor neuros/aPKC
on Monday, March 14th, 2022 11:40 | by Andreas Ehweiner
Overlap of FoxP (red) with aPKC or 3 motor neuro lines (D42, C380, OK6) (green)
aPKC
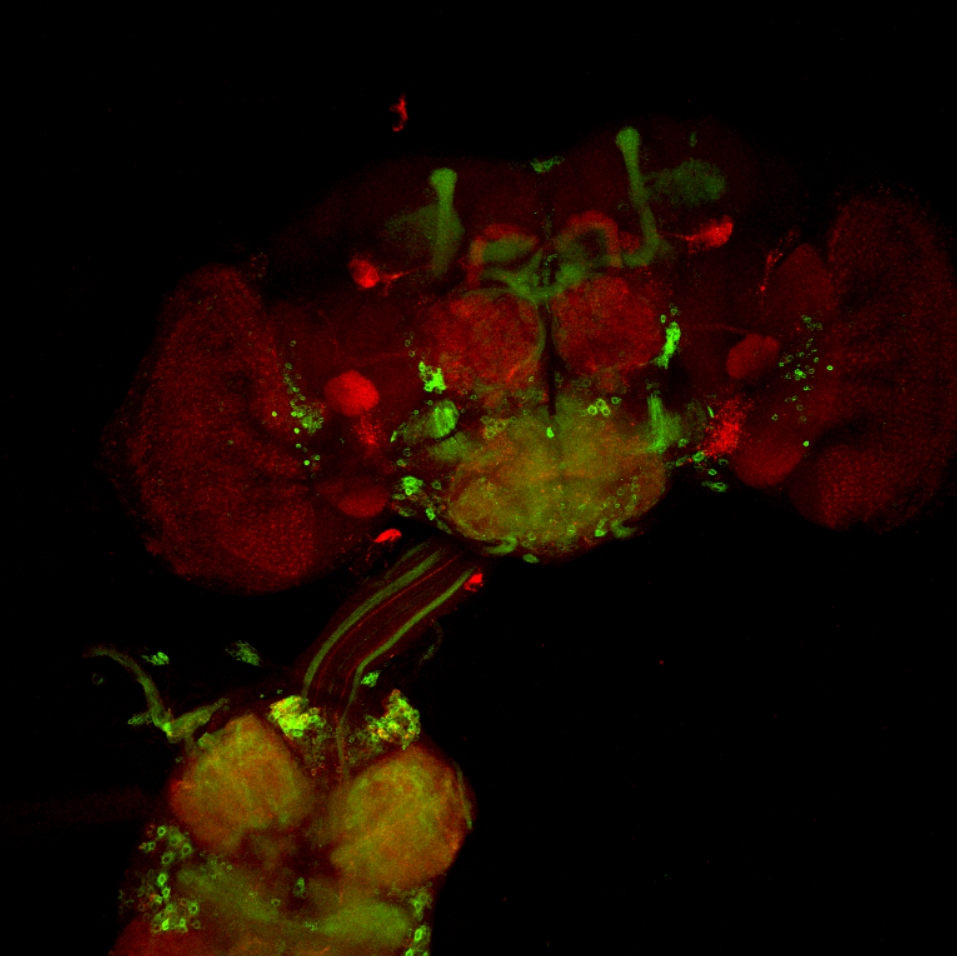
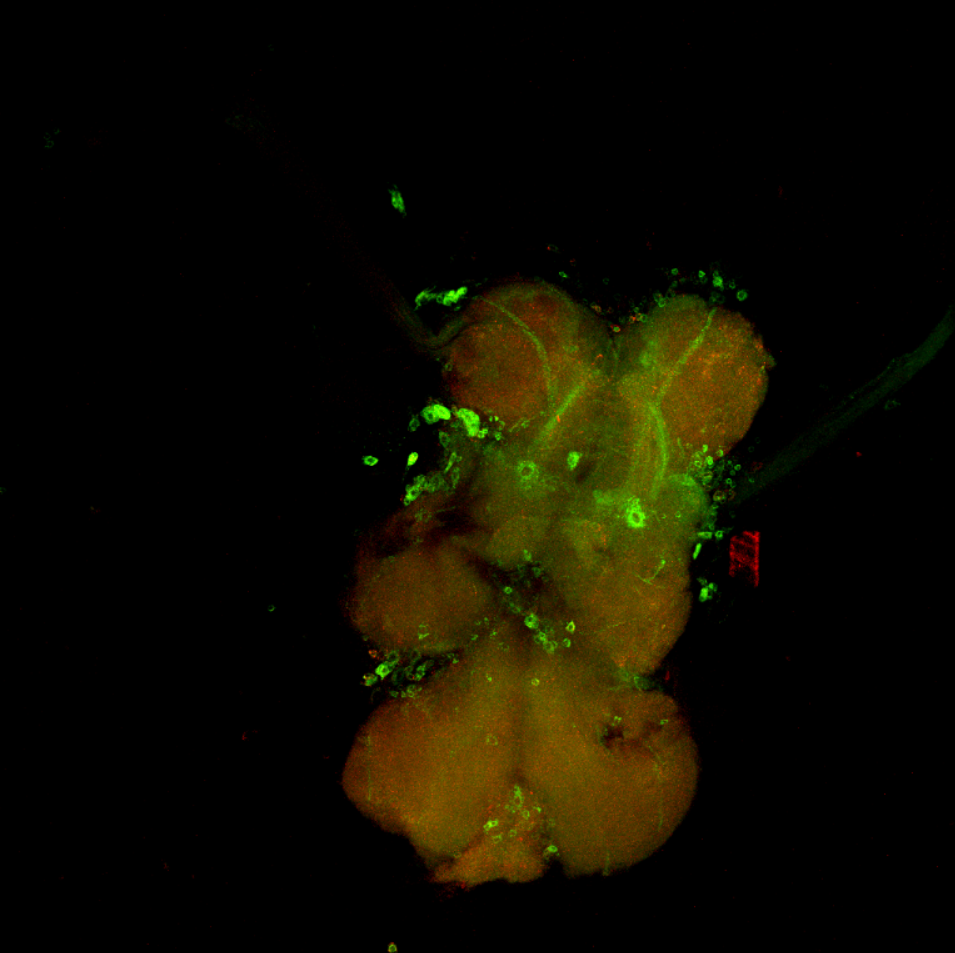
D42
C380
OK6
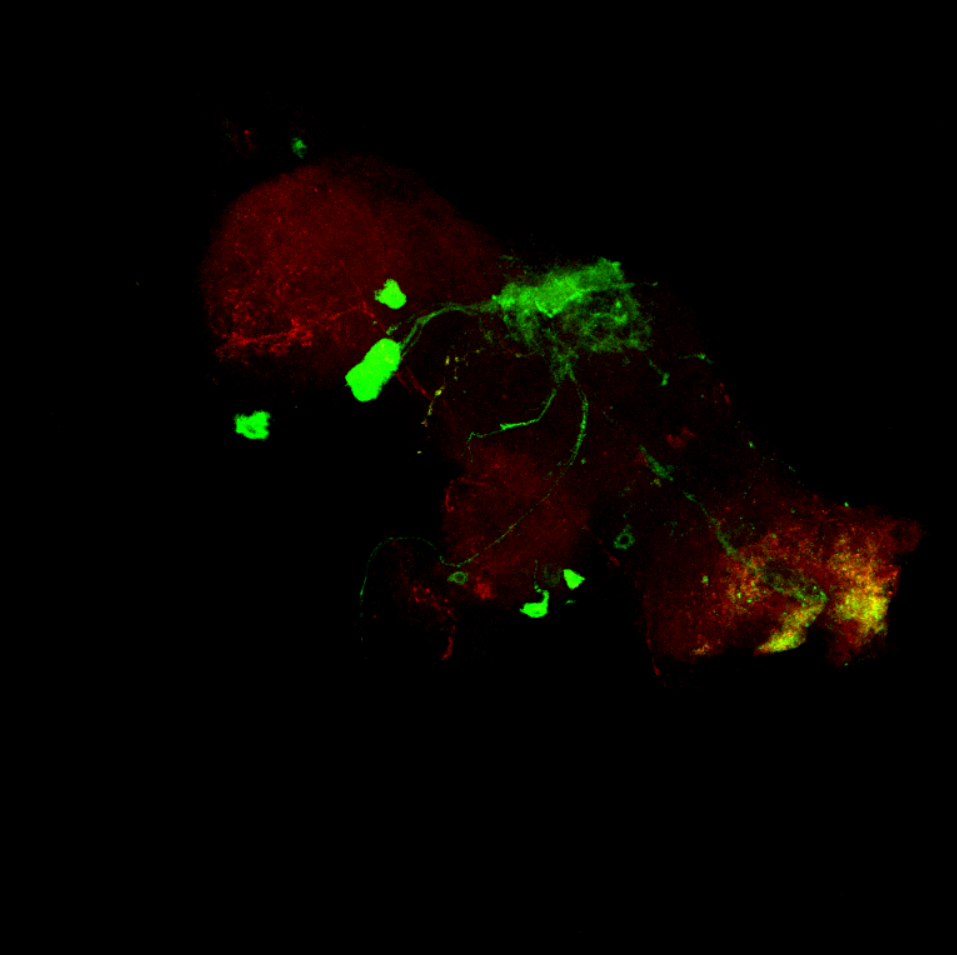
Category: Anatomy, Foxp, PKC, PKC_localisation, Uncategorized | No Comments
Course
on Wednesday, February 16th, 2022 11:29 | by Björn Brembs
https://docs.google.com/spreadsheets/d/1kK-fg84ze-eEPgcxc1-ABjI4WqPdtKlfucU9zZ5TLt8/edit?usp=sharing
Category: Uncategorized | No Comments
Blocking with tetoxE
on Monday, December 20th, 2021 1:37 | by Andreas Ehweiner
tetoxE was crossed to different Gal4-lines. 20A02 was tested separately before, 52B10, 55G08 and 64H04 were tested in parallel.
PB expression: 52B10, 55G08
EB expression: 20A02, 64H04
Previous experiment (Posted 30.08.21):
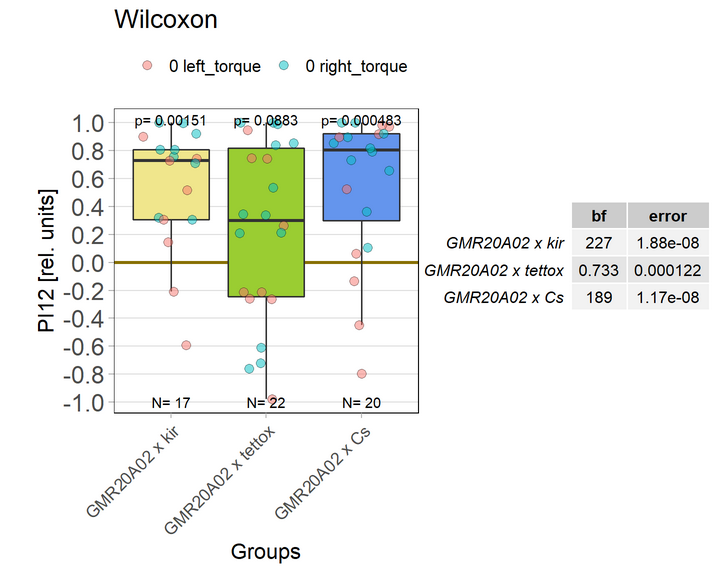
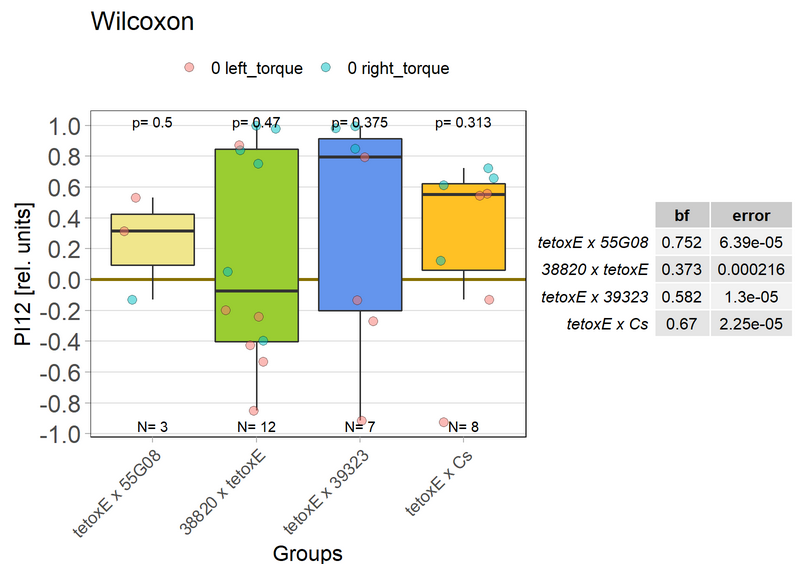
New test
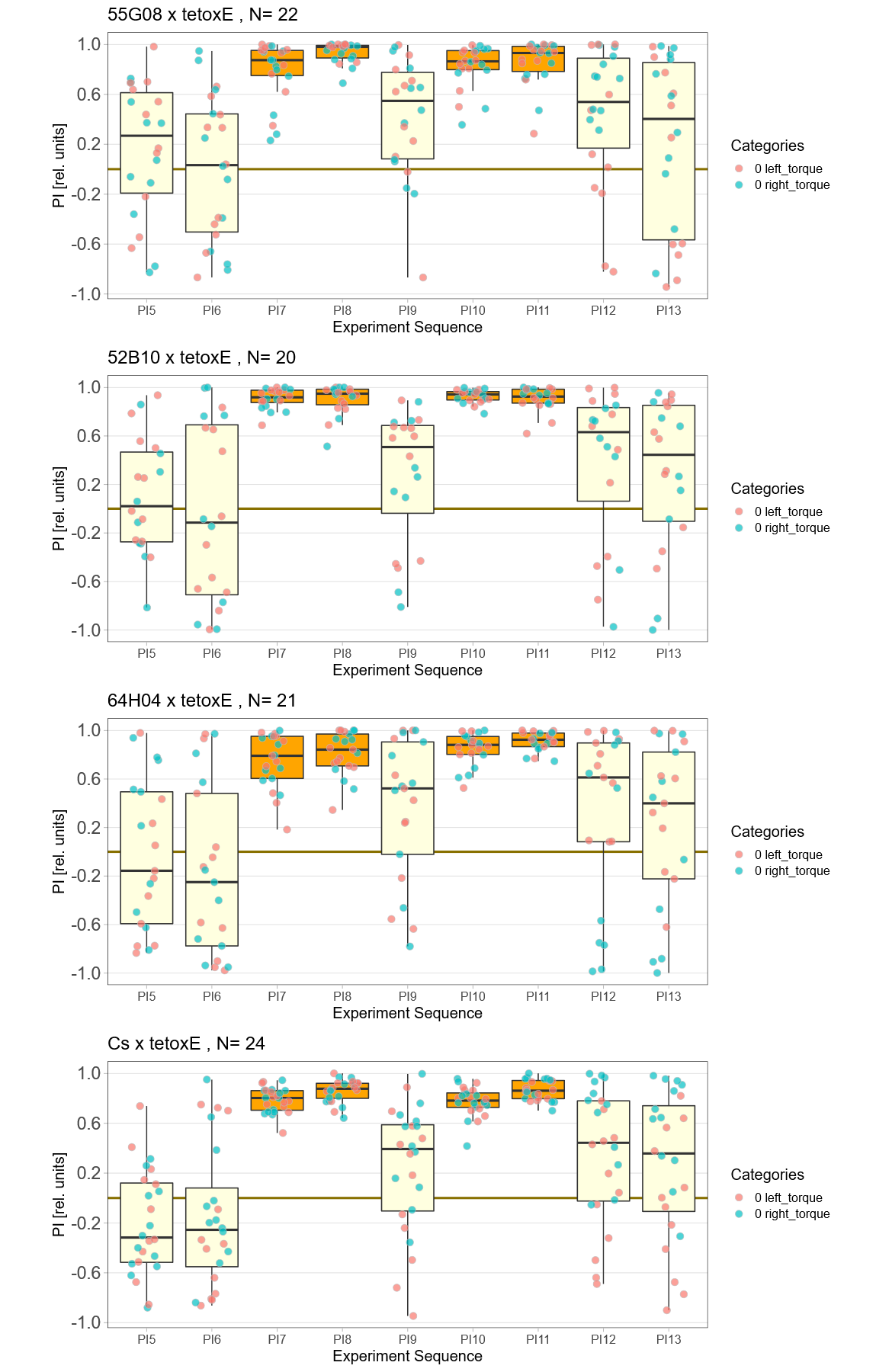
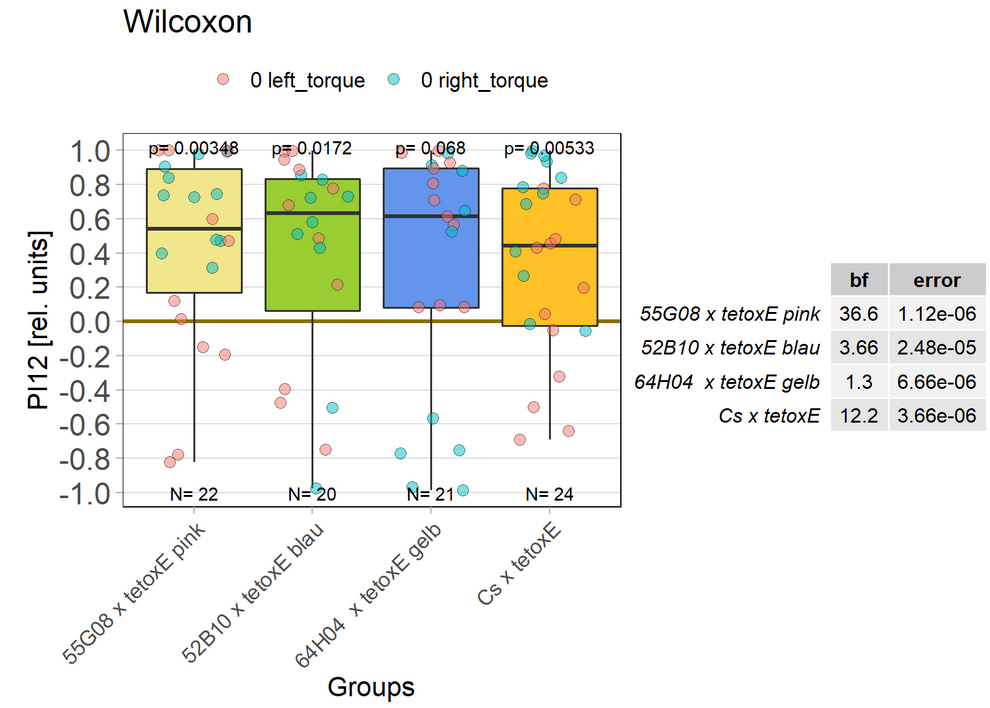
EB: 64H04
Category: flight, Memory, Operant learning, operant self-learning, Uncategorized | No Comments
T-Maze experiment with red light
on Monday, December 13th, 2021 12:33 | by Parva Nasimi

T-Maze experiment with Red light:
Graph shows “Mean Performance Index” for NorpA-UAS-Chrimson/Gr28bd, NorpA-UAS-Chrimson/0273, NorpA-UAS-Chrimson/38764, NorpA-UAS-Chrimson/66555, NorpA-UAS-Chrimson/66010, NorpA-UAS-Chrimson/66009 (ATR-treated foods for 2days) and Controls (NorpA-UAS-Chrimson/Gr28bd, without ATR).
Category: Uncategorized | No Comments
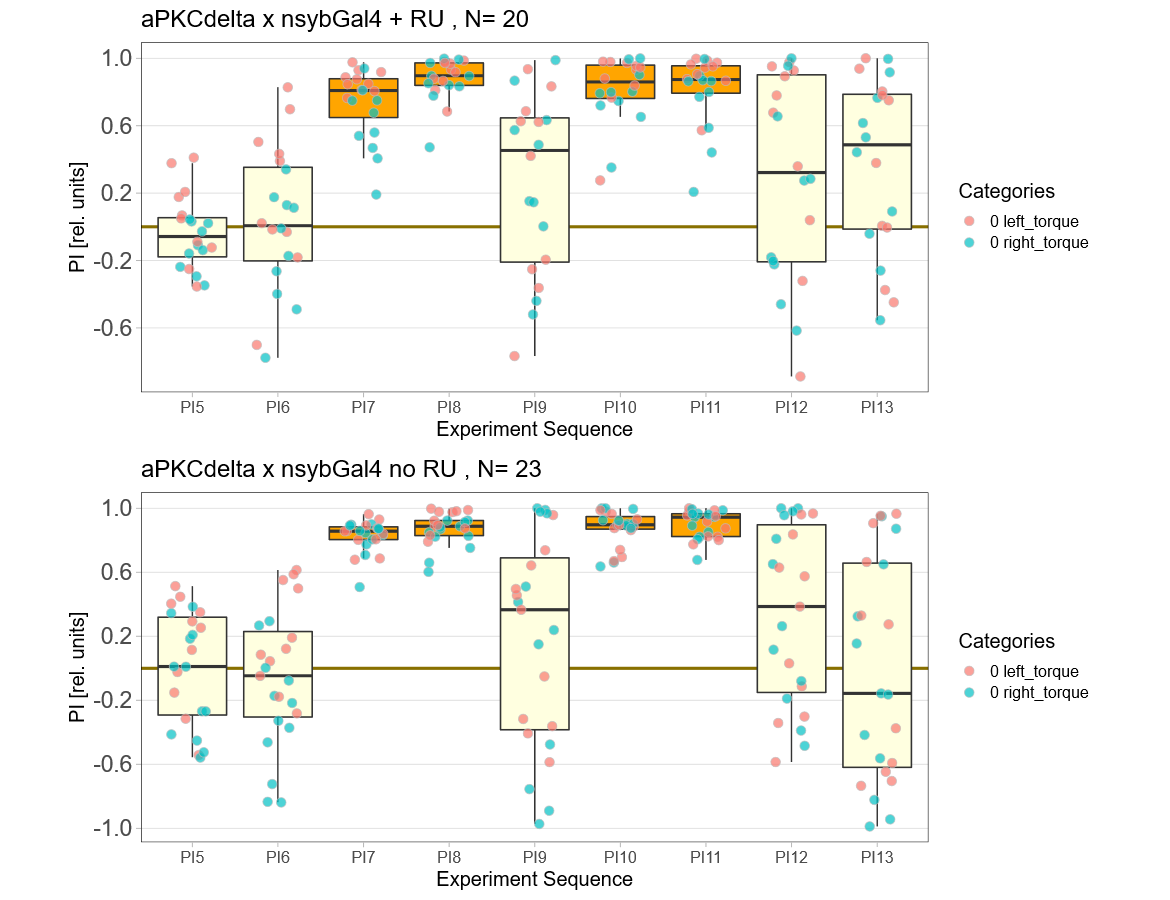
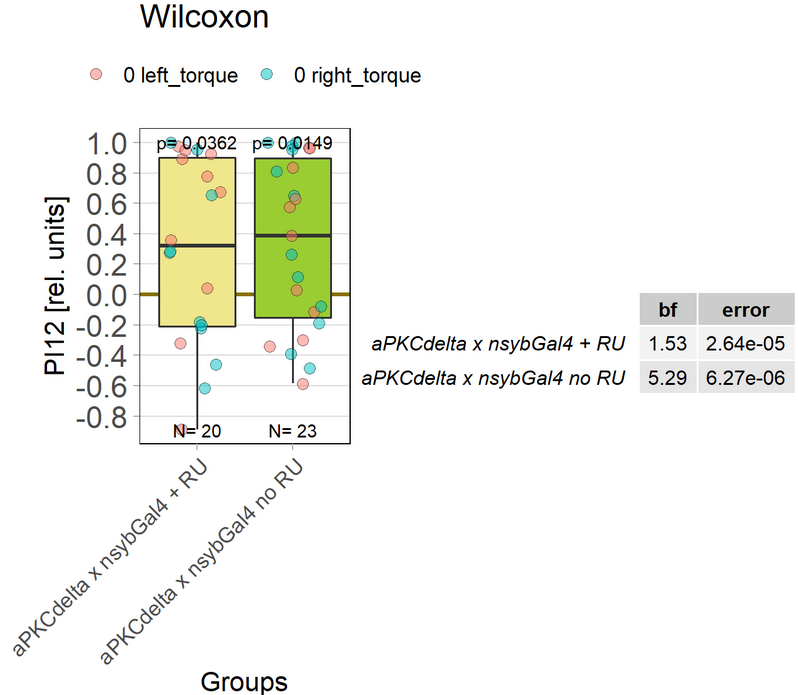
aPKCdelta adult expression
on Monday, December 13th, 2021 9:27 | by Andreas Ehweiner
Expressing aPKCdelta in all neurons in the adult fly.
Category: flight, Memory, Operant learning, operant self-learning, PKC, Uncategorized | No Comments